Climate-Ocean Effects on AYK Chinook Salmon
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Forage Fishes of the Southeastern Bering Sea Conference Proceedings
a OCS Study MMS 87-0017 Forage Fishes of the Southeastern Bering Sea Conference Proceedings 1-1 July 1987 Minerals Management Service Alaska OCS Region OCS Study MMS 87-0017 FORAGE FISHES OF THE SOUTHEASTERN BERING SEA Proceedings of a Conference 4-5 November 1986 Anchorage Hilton Hotel Anchorage, Alaska Prepared f br: U.S. Department of the Interior Minerals Management Service Alaska OCS Region 949 East 36th Avenue, Room 110 Anchorage, Alaska 99508-4302 Under Contract No. 14-12-0001-30297 Logistical Support and Report Preparation By: MBC Applied Environmental Sciences 947 Newhall Street Costa Mesa, California 92627 July 1987 CONTENTS Page ACKNOWLEDGMENTS .............................. iv INTRODUCTION PAPERS Dynamics of the Southeastern Bering Sea Oceanographic Environment - H. Joseph Niebauer .................................. The Bering Sea Ecosystem as a Predation Controlled System - Taivo Laevastu .... Marine Mammals and Forage Fishes in the Southeastern Bering Sea - Kathryn J. Frost and Lloyd Lowry. ............................. Trophic Interactions Between Forage Fish and Seabirds in the Southeastern Bering Sea - Gerald A. Sanger ............................ Demersal Fish Predators of Pelagic Forage Fishes in the Southeastern Bering Sea - M. James Allen ................................ Dynamics of Coastal Salmon in the Southeastern Bering Sea - Donald E. Rogers . Forage Fish Use of Inshore Habitats North of the Alaska Peninsula - Jonathan P. Houghton ................................. Forage Fishes in the Shallow Waters of the North- leut ti an Shelf - Peter Craig ... Population Dynamics of Pacific Herring (Clupea pallasii), Capelin (Mallotus villosus), and Other Coastal Pelagic Fishes in the Eastern Bering Sea - Vidar G. Wespestad The History of Pacific Herring (Clupea pallasii) Fisheries in Alaska - Fritz Funk . Environmental-Dependent Stock-Recruitment Models for Pacific Herring (Clupea pallasii) - Max Stocker. -

Fishery Management Plan for Arctic Grayling Sport Fisheries Along the Nome Road System, 2001–2004
Fishery Management Report No. 02-03 Fishery Management Plan for Arctic Grayling Sport Fisheries along the Nome Road System, 2001–2004 by Fred DeCicco April 2002 Alaska Department of Fish and Game Division of Sport Fish Symbols and Abbreviations The following symbols and abbreviations, and others approved for the Système International d'Unités (SI), are used in Division of Sport Fish Fishery Manuscripts, Fishery Data Series Reports, Fishery Management Reports, and Special Publications without definition. Weights and measures (metric) General Mathematics, statistics, fisheries centimeter cm All commonly accepted e.g., Mr., Mrs., alternate hypothesis HA deciliter dL abbreviations. a.m., p.m., etc. base of natural e gram g All commonly accepted e.g., Dr., Ph.D., logarithm hectare ha professional titles. R.N., etc. catch per unit effort CPUE kilogram kg and & coefficient of variation CV at @ 2 kilometer km common test statistics F, t, , etc. liter L Compass directions: confidence interval C.I. meter m east E correlation coefficient R (multiple) north N metric ton mt correlation coefficient r (simple) milliliter ml south S covariance cov millimeter mm west W degree (angular or ° Copyright temperature) Weights and measures (English) Corporate suffixes: degrees of freedom df cubic feet per second ft3/s Company Co. divided by ÷ or / (in foot ft Corporation Corp. equations) gallon gal Incorporated Inc. equals = inch in Limited Ltd. expected value E mile mi et alii (and other et al. fork length FL ounce oz people) greater than > O pound lb et cetera (and so forth) etc. greater than or equal to quart qt exempli gratia (for e.g., harvest per unit effort HPUE example) yard yd less than < id est (that is) i.e., ? less than or equal to latitude or longitude lat. -

Pamphlet to Accompany Scientific Investigations Map 3131
Bedrock Geologic Map of the Seward Peninsula, Alaska, and Accompanying Conodont Data By Alison B. Till, Julie A. Dumoulin, Melanie B. Werdon, and Heather A. Bleick Pamphlet to accompany Scientific Investigations Map 3131 View of Salmon Lake and the eastern Kigluaik Mountains, central Seward Peninsula 2011 U.S. Department of the Interior U.S. Geological Survey Contents Introduction ....................................................................................................................................................1 Sources of data ....................................................................................................................................1 Components of the map and accompanying materials .................................................................1 Geologic Summary ........................................................................................................................................1 Major geologic components ..............................................................................................................1 York terrane ..................................................................................................................................2 Grantley Harbor Fault Zone and contact between the York terrane and the Nome Complex ..........................................................................................................................3 Nome Complex ............................................................................................................................3 -
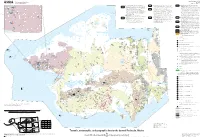
Digital Data for the Preliminary Bedrock Geologic Map of The
Open-File Report 2009-1254 U.S. Department of the Interior Sheet 2 of 2 U.S. Geological Survey Pamphlet accompanies map '02°761 '01°761 '00°761 LIST OF METAMORPHIC-TECTONIC ELEMENTS HIGH GRADE METAMORPHIC AND ASSOCIATED O<t Qs Ol Nome Complex, west-central – Weakly foliated metasedimentary O<l O<l IGNEOUS ROCKS – Amphibolite and granulite-facies O<l O<l Ol Ols Ols YORK TERRANE – Late Proterozoic (?) and Paleozoic sedimentary and unfoliated metaigneous rocks that retain relict primary features; and metasedimentary rocks and minor Late Cretaceous tin-bearing mineral assemblages in mafic rocks formed at pumpellyite-actinolite, metamorphic rocks and associated Cretaceous plutons; penetratively Qs greenschist, and blueschist facies (one locality) deformed metasedimentary and metaigneous schist and gneiss with Oal granites; dominantly carbonate and siliciclastic lithologies, in which Oal primary features are generally retained; fine-grained Nome Complex, eastern – Penetratively deformed and complex metamorphic histories; aluminum-rich lithologies show Ol early development of kyanite-stable mineral assemblages succeeded Ol 17 metasedimentary rocks are weakly foliated. Metamorphic and recrystallized schists with ductile fabrics; protolith packages and Ol Ols 165°00' by sillimanite-stable, lower-pressure assemblages. Lithologies rich in O<p Ols thermal history variable from unit to unit, and generally lower grade metamorphic fabrics identical to Nome Complex in central Seward Oal Qs Peninsula; mineral assemblages in most of the area are characteristic iron and aluminum retain early, relatively high pressure Ktg than Nome Complex. Tin granites intruded in shallow crustal Ols Ol aluminosilicate plus orthoamphibole assemblages (>5kb) that are Cape Espenberg settings. The generally brittle shallow and steeply-dipping structures of greenschist facies, but slightly higher grade assemblages occur in Ol the vicinity of Kiwalik Mountain. -

The PAYSTREAK Volume 12, No
The PAYSTREAK Volume 12, No. 2, Fall 2010 The Newsletter of the Alaska Mining Hall of Fame Foundation (AMHF) In this issue: AMHF New Inductees Page 1 Induction Ceremony Program Page 2 Introduction and Acknowledgements Page 3 Previous AMHF Inductees Pages 4 - 10 New Inductee Biographies Pages 10 - 23 Distinguished Alaskans Aid Foundation Page 24 Alaska Mining Hall of Fame Directors and Officers Page 24 Alaska Mining Hall of Fame Foundation New Inductees AMHF Honors Pioneers Important to the Seward Peninsula Gold Dredging Industry Nicholas B. and Evinda S. Tweet: It is difficult to name any couple in Alaska mining history that has had more longevity and perseverance than Nicholas B. and Evinda S. Tweet. In marriage, they formed a team that created a remarkably stable, family-owned firm, N.B. Tweet and Sons, which has mined gold in Alaska for 110 years. Nicholas and Evinda mined and lived in several placer mining camps, worked graphite claims, operated gold dredges, and inspired their descendants to continue the placer mining lifestyle. Both Evinda and Nick died at the family mining camp, near Taylor, north of Nome. In 2010, N.B. Tweet and Sons operated the only bucketline stacker gold dredge in North America. Carl S. (left) and Walter A. (right) Glavinovich: Two Croatian (Yugoslavian)-born brothers who, collectively, devoted more than one-hundred years of their lives to the prospecting, deciphering, drilling, thawing, and dredging of the Nome, Alaska, placer gold fields. Most of these years were in the service of one company, the U.S. Smelting, Refining, and Mining Company (USSR&M) or its direct affiliate, the Hammon Consolidated Gold Fields. -

Report of Survey and Inventory Activities, Walrus Studies
ll'tl-LKl"l'-'llul'\ L.r-rt1,..Lr-. I <; E SECTILI'• Al.if <; G JlJM:AlJ ALASKA DEPARTMENT OF FISH AND GAME J UN E A U, AL AS KA STATE OF ALASKA William A. Egan, Governor DEPARTMENT OF FISH AND GAME James W. Brooks, Commissioner DIVISION OF GAME Frank Jones, Director RE P 0 RT 0 F S uRV E Y & I NV E NT 0 RY AC T I VI T I ES WALRUS STUDIES by John J. Burns Volume I Federal Aid in Wildlife Restoration Project W-17-3, Job 8.0 and Project W-17-4, Job 8.0 (1st half) Persons are free to use material in these reports for educational or informational purposes. However, since most reports treat only part of continuing studies, persons intending to use this material in scientific publications should obtain prior permission from the Department of Fish and Game. In all cases tentative conclusions should be identified as such in quotation, and due credit would be appreciated. (Printed January, 197J) SURVEY-INVENTORY PROGRESS REPORT State: Alaska Cooperators: Alexander Akeya, Savoonga; Rae Baxter, Alaska Department of Fish and Game; Carl Grauvogel, Alaska Department of Fish and Game; Edward Muktoyuk, Alaska Department of Fish and Game; Robert Pegau, Alaska Department of Fish and Game; and Vernon Slwooko, Gambell Project Nos • : W-17-3 & Project Title: Marine Mammal S&I W-17-4 Job No.: Job Title: Walrus Studies Period Covered: July l, 1970 to December 31, 1971. SUMMARY The 1971 harvest of walruses in Alaska was 1915 animals. Of these, 1592 (83%) were bulls, 254 (13%) cows and 69 (4%) calves of either sex. -

17 R.-..Ry 19" OCS Study MMS 88-0092
OCISt.., "'1~2 Ecologic.1 Allociue. SYII'tUsIS 0' ~c. (I( 1'B IPnCfS OP MOISE AlII) DIsmuA1K2 a, IIUc. IIADLOft m.::IIIDA1'IONS or lUIS SIA PI.-IPms fr •• LGL Muke ••••• rda Aaeoc:iat_, Inc •• 505 "-t IIortbera Lllbta .1••••,"Sait. 201 ABdaonp, AlMke 99503 for u.s. tIl_rala •••••••••• Seni.ce Al_1taa o.t.r CoIItlM11tal Shelf legion U.S. u.,c. of Iat.dor ••• 603, ,., EMt 36tla A.-... A8eb0ra•• , Aluke 99501 Coatraet _. 14-12-00CU-30361 LGL •••••• 'U 821 17 r.-..ry 19" OCS Study MMS 88-0092 StAllUIS OWIUOlMUc. 011'DB &IIBCfI ,. 11010 AlII) DIS'ftJU8CB 011llAJoa IWJLOU'r COIICIII'DArIa. OW101. SB&PIDU&DI by S.R. Johnson J.J. Burnsl C.I. Malme2 R.A. Davis LGL Alaska Research Associatel, Inc. 505 West Northern Lights Blv~., Suite 201 Anchorage, Alaska 99503 for u.S. Minerals Management Service Alaskan Outer Continental Shelf Region U.S. Dept. of Interior Room 603, 949 East 36th Avenue Anchorage, Alaska 99508 Contract no. 14-12-0001-30361 LGL Rep. No. TA 828 17 February 1989 The opinionl, findings, conclusions, or recolmBendations expressed in this report are those of the authors and do not necessarily reflect the views of the U.S. Dept. of the Interior. nor does mention of trade names or commercial products constitute endorsement or recommendation for use by the Federal Government. 1 Living Resources Inc., Fairbanks, AK 2 BBN Systems and Technologies Corporation, Cambridge, MA Table of Contents ii 'UIU or cc»mll UBLBor cowmll ii AIS'lIAC'f • · . vi Inter-site Population Sensitivity Index (IPSI) vi Norton Basin Planning Area • • vii St. -

Distribution and Migration and Status of Pacific Herring
AYK Herring Report No.lZ ~ DISTRIBUTION !ND MIGRATION AND STATUS OF PACIFIC HERRING by Vidar G. Wespestad* and Louis H. Barton** *Northwest and Alaska Fisheries Center National Marine Fisheries Service National Oce~ic and Atmospheric Administration 2725 Montlake Boulevard East Seattle, Washington 98112 **Alaska Dept. Fish and Game 333 Raspberry Road Anchorage, Alaska 99502 December 1979 .. .. .. .. .. DISTRIBUTION AND MIGRATION AND STATUS OF PACIFIC HERRING Vidar G. Wespestad Northwest and Alaska Fisheries Center 2725 Montlake Blvd. E. Seattle, Washington 98112 Louis H. Barton Alaska Dept. Fish and Game 333 Raspberry Road Anchorage, Alaska 99502 ABSTRACT Pacific herring are an important part of the Bering Sea food web and fi;>rm the basis of a major connnercial fishery. Until re cently Japan and the U.S.S.R. have been major exploiters of herring. Catch peaked in the early 1970 1 s at 145,579 mt, and then declined in response to overfishing and poor recruitment. Recently herring abundance has increased, and the United States has become the dom inant exploiter of herring. Most herring are harvested in coastal waters during the spawn ing period which commences in late April/mid-May along the Alaska Peninsula and Bristol Bay and progressively later to the north. Spawning occurs at temperatures of 5-12 C and the time of spawning is related to winter water temperatures with early spawning in warm years and 1ate spawning in co1d years. Ouri ng spawning eggs are 166. deposited on vegetation in the intertidal zone of shallow bays and rocky headlands. Eggs hatch in 2-3 weeks as planktonic larvae and metamorphose to juveniles after 6-10 weeks. -

Alaska OCS Socioeconomic Studies Program
. i, WUOE (ixlw Technical Report Number 54 Volume 1 -, Alaska OCS Socioeconomic Studies Program Sponsor: Bureau of Land Management — Alaska Outer ‘ Bering–Normn Pwdeum Development Scenarios Sociocultural Syst~ms Analysis The United States Department of the Interior was designated by the Outer Continental Shelf (OCS) Lands Act of 1953 to carry out the majority of the Act’s provisions for administering the mineral leasing and develop- ment of offshore areas of the United States under federal jurisdiction. Within the Department, the Bureau of Land Management (ELM) has the responsibility to meet requirements of the National Environmental Policy Act of 1969 (NEPA) as well as other legislation and regulations dealing with the effects of offshore development. In Alaska, unique cultural differences and climatic conditions create a need for developing addi- tional socioeconomic and environmental information to improve OCS deci- sion making at all governmental levels. In fulfillment of its federal responsibilities and with an awareness of these additional information needs, the BLM has initiated several investigative programs, one of which is the Alaska OCS Socioeconomic Studies Program (SESP). The Alaska OCS Socioeconomic Studies Program is a multi-year research effort which attempts to predict and evaluate the effects of Alaska OCS Petroleum Development upon the physical, social, and economic environ- ments within the state. The overall methodology is divided into three broad research components. The first component identifies an alterna- tive set of assumptions regarding the location, the nature, and the timing of future petroleum events and related activities. In this component, the program takes into account the particular needs of the petroleum industry and projects the human, technological, economic, and environmental offshore and onshore development requirements of the regional petroleum industry. -

Seward Peninsula RAC Transcript Volume 1 Winter 2020
SEWARD PENINSULA SUBSISTENCE RAC MEETING 3/11/2020 SEWARD PENINSULA RAC MEETING 1 SEWARD PENINSULA FEDERAL SUBSISTENCE REGIONAL ADVISORY COUNCIL MEETING PUBLIC MEETING VOLUME I Nome Mini-Convention Center Nome, Alaska March 11, 2020 9:07 a.m. Members Present: Tom Gray, Acting Chairman Deahl Katchatag Ronald Kirk Lloyd Kiyutelluk Leland Oyoumick Charles Saccheus Elmer Seetot Regional Council Coordinator -Tom Kron (Acting) Karen Deatherage/phone Recorded and transcribed by: Computer Matrix Court Reporters, LLC 135 Christensen Drive, Suite 2 Anchorage, AK 99501 907-227-5312; [email protected] Computer Matrix, LLC Phone: 907-243-0668 135 Christensen Dr., Ste. 2., Anch. AK 99501 Fax: 907-243-1473 SEWARD PENINSULA SUBSISTENCE RAC MEETING 3/11/2020 SEWARD PENINSULA RAC MEETING 1 Page 2 1 P R O C E E D I N G S 2 3 (Nome, Alaska - 3/11/2020) 4 5 (On record) 6 7 ACTING CHAIR GRAY: If I could get 8 everybody to stand I'd appreciate it. Take your hat 9 off. So I'm going to give the invocation. 10 11 (Invocation) 12 13 ACTING CHAIR GRAY: Thank you. Okay, 14 so I'm going to call the meeting to order so we're 15 official. And I get to be the guy running the meeting. 16 Louis is -- he called me from Ruby yesterday saying 17 they're stuck -- they're not stuck, they're actually 18 moving again but him and his brother and nieces and so 19 on and so forth are driving snowmachines to Nome from 20 somewhere, and it's taken longer than they expected. -

Of Fish and Wildlife
Alasha Habitat Management Guide Life Histories and Habitat Requirements of Fish and Wildlife Produced b'y State of Alasha Department of Fish and Game Division of Habitat Iuneau, Alasha 1986 @ Tgge BY TTIE AI.ASKA DEPARTMENT OF FISH AI{D GA}IE Contents Achnowtedgemena v i i Introduction 3 0verview of Habitat l{anagement Guides Project 3 Background 3 Purpose 3 Appl ications 4 Statewide volumes 6 Regional volumes 9 0rganization and Use of this Volume 10 Background 10 Organization 10 Species selection criteria 10 Regional Overview Surmaries 11 Southwest Region 11 Southcentral Region 13 Arctic Region 15 hlestern and Interior regions L7 ,lf{atnmals Mari ne Manmal s --BEfiEti-Thale zr Bowhead whale 39 Harbor seal 55 Northern fur seal 63 Pacific walrus 7L Polar bear 89 Ringed seal 107 Sea otter 119 Steller sea lion 131 ffiTerrestrial Mamrnals Brown bear 149 Caribou 165 Dall sheep 175 El k 187 Moose 193 Si tka bl ack-tai'led deer 209 1,1,L Contents (continued) Birds Bald Eagle 229 Dabbl ing ducks 241 Diving ducks 255 Geese 27L Seabirds 29L Trumpeter swan 301 Tundra swan 309 Fish Freshwater/Anadromous Fi sh en 3L7 Arctic aray'ling 339 Broad whitefish 361 Burbot 377 Humpback whitefish 389 Lake trout 399 Least cisco 413 Northern pike 425 Rainbow trout/steelhead 439 Sa I mon Chum 457 Chinook 481 Coho 501 Pink 519 Sockeye 537 Sheefish 555 l.lari ne Fi sh cod 57L --ffiiTCapel in 581 Pacific cod 591 Pacific hal ibut 599 Pacific herring 607 Pacific ocean perch 62L Sablefish 629 Saffron cod 637 Starry flounder 649 t{al leye pol lock 657 Yelloweye rockfish 667 Yel lowfin sole 673 Contents (continued) Shel I fi sh crab 683 King crab 691 -ungenessRazor clam 709 Shrimp 7L5 Tanner crab 727 Appendi ces A. -

Investigations of Belukha Whales in Coastal Waters
INVESTIGATIONS OF BELUKHA WHALES IN COASTAL WATERS OF WESTERN AND NORTHERN ALASKA I. DISTRIBUTION, ABUNDANCE, AND MOVEMENTS by Glenn A. Seaman, Kathryn J. Frost, and Lloyd F. Lowry Alaska Department of Fish and Game 1300 College Road Fairbanks, Alaska 99701 Final Report Outer Continental Shelf Environmental Assessment Program Research Unit 612 November 1986 153 TABLE OF CONTENTS Section Page LIST OF FIGURES . 157 LIST OF TABLES . 159 sumRY . 161 ACKNOWLEDGEMENTS . 162 WORLD DISTRIBUTION . 163 GENERAL 1 ISTRIBUTION IN ALASKA . 165 SEASONAL DISTRIBUTION IN ALASKA. 165 REGIONAL DISTRIBUTION AND ABUNDANCE. 173 Nor” h Aleutian Basin. 173 Saint Matthew-Hall Basin. s . 180 Saint George Basin. 184 Navarin Basin . 186 Norton Basin. 187 Hope Basin. 191 Barrow Arch . 196 Diapir Field. 205 DISCUSSION AND CONCLUSIONS . 208 LITERATURE CITED . 212 LIST OF PERSONAL COMMUNICANTS. 219 155 LIST OF FIGURES Figure 1. Current world distribution of belukha whales, not including extralimital occurrences . Figure 2. Map of the Bering, Chuk&i, and Beaufort seas, showing major locations mentioned in text. Figure 3. Distribution of belukha whales in January and February . Figure 4. Distribution of belukha whales in March and April. Figure 5. Distribution of belukha whales in May and June . Figure 6. Distribution of belukha whales in July and August. Figure 7. Distribution of belukha whales in September and October. Figure 8. Distribution of belukha whales in November and Deeember. Figure 9. Map of the North Aleutian Basin showing locations mentioned in text. Figure 10. Map of the Saint Matthew-Hall Basin showing locations mentionedintext. Figure 11. Map of the Saint George and Navarin basins showing locations mentionedintext.