NEUROANATOMY of the VISUAL PATHWAYS Magrane Basic Science Course June 2018
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
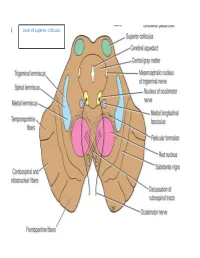
Level of Superior Colliculus ° Edinger- Westphal Nucleus ° Pretectal Nucleus: Close to the Lateral Part of the Superior Colliculus
Level of superior colliculus ° Edinger- Westphal nucleus ° pretectal nucleus: close to the lateral part of the superior colliculus. Red nucleus • Rounded mass of gray matter • Situated bt cerebral aqueduct and substantia nigra • Reddish blue(vascularity & iron containing pigment) • Afferents from: cerebral cortex,cerebellum,substa ntia nigra, thalamic nuclei, spinal cord • Efferent to: spinal cord, reticular formation. thalamus and substantia nigra • involved in motor coordination. Crus cerebri • Corticospinal & corticonuclear fibers (middle) • Frontopontine fibers (medial) • Temporopontine fibers (lateral) these descending tracts connect the cerebral cortex with spinal cord, cranial nerves nuclei, pons & cerebellum Level at superior colliculus ° Superior colliculus ° Occulomotor nucleus (posterior to MLF) ° Occulomotor n emerges through red nucleus ° Edinger-Westphal nucleus ° pretectal nucleus: close to the lateral part of the superior colliculus. ° MLF ° Medial , trigeminal, spinal leminiscus ( no lateral leminiscus) ° Red nucleus ° Substantia nigra ° Crus cerebri ° RF Substantia nigra ° Large motor nucleus ° is a brain structure located in the midbrain ° plays an important role in reward, addiction, and movement. ° Substantia nigra is Latin for "black substance" due to high levels of melanin ° has connections with basal ganglia ,cerebral cortex ° Concerned with muscle tone ° Parkinson's disease is caused by the death of neurons in the substantia nigra Oculomotor Nerve (III) • Main oculomotor nucleus • Accessory parasympathetic nucleus -

Dissection and Exposure of the Whole Course of Deep Nerves in Human Head Specimens After Decalcification
Hindawi Publishing Corporation International Journal of Otolaryngology Volume 2012, Article ID 418650, 7 pages doi:10.1155/2012/418650 Research Article Dissection and Exposure of the Whole Course of Deep Nerves in Human Head Specimens after Decalcification Longping Liu, Robin Arnold, and Marcus Robinson Discipline of Anatomy and Histology, University of Sydney, Anderson Stuart Building F13, Sydney, NSW 2006, Australia Correspondence should be addressed to Marcus Robinson, [email protected] Received 29 July 2011; Revised 10 November 2011; Accepted 12 December 2011 AcademicEditor:R.L.Doty Copyright © 2012 Longping Liu et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The whole course of the chorda tympani nerve, nerve of pterygoid canal, and facial nerves and their relationships with surrounding structures are complex. After reviewing the literature, it was found that details of the whole course of these deep nerves are rarely reported and specimens displaying these nerves are rarely seen in the dissecting room, anatomical museum, or atlases. Dissections were performed on 16 decalcified human head specimens, exposing the chorda tympani and the nerve connection between the geniculate and pterygopalatine ganglia. Measurements of nerve lengths, branching distances, and ganglia size were taken. The chorda tympani is a very fine nerve (0.44 mm in diameter within the tympanic cavity) and approximately 54 mm in length. The mean length of the facial nerve from opening of internal acoustic meatus to stylomastoid foramen was 52.5 mm. -

Dissection and Exposure of the Whole Course of Deep Nerves in Human Head Specimens After Decalcification
Hindawi Publishing Corporation International Journal of Otolaryngology Volume 2012, Article ID 418650, 7 pages doi:10.1155/2012/418650 Research Article Dissection and Exposure of the Whole Course of Deep Nerves in Human Head Specimens after Decalcification Longping Liu, Robin Arnold, and Marcus Robinson Discipline of Anatomy and Histology, University of Sydney, Anderson Stuart Building F13, Sydney, NSW 2006, Australia Correspondence should be addressed to Marcus Robinson, [email protected] Received 29 July 2011; Revised 10 November 2011; Accepted 12 December 2011 AcademicEditor:R.L.Doty Copyright © 2012 Longping Liu et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The whole course of the chorda tympani nerve, nerve of pterygoid canal, and facial nerves and their relationships with surrounding structures are complex. After reviewing the literature, it was found that details of the whole course of these deep nerves are rarely reported and specimens displaying these nerves are rarely seen in the dissecting room, anatomical museum, or atlases. Dissections were performed on 16 decalcified human head specimens, exposing the chorda tympani and the nerve connection between the geniculate and pterygopalatine ganglia. Measurements of nerve lengths, branching distances, and ganglia size were taken. The chorda tympani is a very fine nerve (0.44 mm in diameter within the tympanic cavity) and approximately 54 mm in length. The mean length of the facial nerve from opening of internal acoustic meatus to stylomastoid foramen was 52.5 mm. -

Clinical Anatomy of Greater Petrosal Nerve and Its Surgical Importance
[Downloaded free from http://www.indianjotol.org on Monday, August 18, 2014, IP: 218.241.189.21] || Click here to download free Android application for this journal ORIGINAL ARTICLE Clinical anatomy of greater petrosal nerve and its surgical importance Prashant E Natekar, Fatima M De Souza Department of Anatomy, Goa Medical College, Bambolim, Goa, India Background: Surgical approach towards greater petrosal nerve has to be done with caution as many surgeons ABSTRACT are unfamiliar with the anatomy of the facial nerve. The anatomical landmarks selected must be reliable and above all easy to identify for identification of the greater petrosal nerve so as to avoid injury to the structures in the middle cranial fossa. Observation and Results: The present study is carried out on 100 temporal bones by examining the following measurements of the right and the left sides a) length of the hiatus for grater petrosal superficial nerve b) distance from superior petrosal sinus c) distance from lateral margin of middle cranial fossa d) arcuate eminence and e) distance from exit to the foramen ovale. Conclusion: The anatomical landmarks selected must be reliable and above all easy to identify. Bony structures are more suitable than soft tissue or cartilaginous landmarks because of their rigid and reliable location. These anatomical landmarks will definitely help the surgeon while performing vidian nerve neurectomy and also the anatomical relationship of the facial nerve in temporal bone. The middle fossa approach involves a temporal craniotomy in cases of perineural spread of adenoid cystic carcinomas hence these anatomical landmarks will serve as useful guide for the surgeons and radiologists. -

NASAL ANATOMY Elena Rizzo Riera R1 ORL HUSE NASAL ANATOMY
NASAL ANATOMY Elena Rizzo Riera R1 ORL HUSE NASAL ANATOMY The nose is a highly contoured pyramidal structure situated centrally in the face and it is composed by: ü Skin ü Mucosa ü Bone ü Cartilage ü Supporting tissue Topographic analysis 1. EXTERNAL NASAL ANATOMY § Skin § Soft tissue § Muscles § Blood vessels § Nerves ² Understanding variations in skin thickness is an essential aspect of reconstructive nasal surgery. ² Familiarity with blood supplyà local flaps. Individuality SKIN Aesthetic regions Thinner Thicker Ø Dorsum Ø Radix Ø Nostril margins Ø Nasal tip Ø Columella Ø Alae Surgical implications Surgical elevation of the nasal skin should be done in the plane just superficial to the underlying bony and cartilaginous nasal skeleton to prevent injury to the blood supply and to the nasal muscles. Excessive damage to the nasal muscles causes unwanted immobility of the nose during facial expression, so called mummified nose. SUBCUTANEOUS LAYER § Superficial fatty panniculus Adipose tissue and vertical fibres between deep dermis and fibromuscular layer. § Fibromuscular layer Nasal musculature and nasal SMAS § Deep fatty layer Contains the major superficial blood vessels and nerves. No fibrous fibres. § Periosteum/ perichondrium Provide nutrient blood flow to the nasal bones and cartilage MUSCLES § Greatest concentration of musclesàjunction of upper lateral and alar cartilages (muscular dilation and stenting of nasal valve). § Innervation: zygomaticotemporal branch of the facial nerve § Elevator muscles § Depressor muscles § Compressor -

Lecture 7 Anatomy the PTERYGOPALATINE FOSSA
د.احمد فاضل القيسي Lecture 7 Anatomy THE PTERYGOPALATINE FOSSA The pterygopalatine fossa lies beneath the posterior surface of the maxilla and the pterygoid process of the sphenoid bone. The pterygopalatine fossa contains the maxillary nerve, the maxillary artery (third part) and the pterygopalatine parasympathetic ganglion. Boundaries Anteriorly: posterior surface of maxilla. Posteriorly: anterior margin of pterygoid process below and greater wing of sphenoid above. Medially: perpendicular plate of palatine bone. Superiorly: greater wing of sphenoid. Laterally: communicates with infratemporal fossa through pterygomaxillary fissure Communications and openings: 1. The pterygomaxillary fissure: transmits the maxillary artery from the infratemporal fossa, the posterior superior alveolar branches of the maxillary division of the trigeminal nerve and the sphenopalatine veins. 2. The inferior orbital fissure: transmits the infraorbital and zygomatic branches of the maxillary nerve, the orbital branches of the pterygopalatine ganglion and the infraorbital vessels. 3. The foramen rotundum from the middle cranial fossa, occupying the greater wing of the sphenoid bone and transmit the maxillary division of the trigeminal nerve 4. The pterygoid canal from the region of the foramen lacerum at the base of the skull. The pterygoid canal transmits the greater petrosal and deep petrosal nerves (which combine to form the nerve of the pterygoid canal) and an accompanying artery derived from the maxillary artery. 5. The sphenopalatine foramen lying high up on the medial wall of the fossa.This foramen communicates with the lateral wall of the nasal cavity. It transmits the nasopalatine and posterior superior nasal nerves (from the pterygopalatine ganglion) and the sphenopalatine vessels. 6. The opening of a palatine canal found at the base of the fossa. -

The Nose Embryology the Nose Developed from the Frontonasal Process
The Nose Embryology The nose developed from the frontonasal process. Anatomy The nose is formed of external nose and nasal cavity (A) The external nose Pyramidal in shape shows: The skeleton is formed of: 1- Bony upper 1/3: - Two nasal bones. - Nasal (frontal) process of maxilla. - Nasal process of frontal bone. 2- Cartilaginous lower 2/3: - Upper lateral cartilage. - Lower lateral cartilage. - Septal cartilage. (B) Nasal cavity Each nasal cavity has 2 openings and 4 walls. 1- Anterior nares or nostrils: separated by the columella. 2- Posterior nares or choanae: open into nasopharynx. 3- Roof: cribriform plate of ethmoid bone, frontal bone ant. 4- Floor: hard palate 5- Medial wall: Septum - Vertical plate of ethmoid: postero superior. - Vomer bone postero inferior. - Quadrangular Septal cartilage - Maxillary crest inferiorly. 6- Lateral wall: formed by medical wall of maxilla(ethmoidal labrynth) , vertical plate of palatine bone, and (inner aspect of nasal bone, frontal process of maxilla, lacrimal bone) it shows: A- 3 bony projections: superior, middle, and inferior conchae or turbinates. B- Each turbinate covers a recess called meatus, there are superior, middle, and inferior meati, in addition to spheno-ethmoidal recess above the superior turbinate. - Inferior meatus drains nasolacrimal duct(site of I.N.A) - Middle meatus drains. 1 Anterior ethmoids via bulla ethmoidalis. Frontal sinus via ant. part of hiatus semilunaris. Maxillary sinus via postero inf. Part or hiatus semi lunaris. - Superior meatus drains: posterior ethmoids. - Spheno- ethmoidal recess drains sphenoid sinus. Blood supply (A) Arterial External nose Nasal cavity - Facial → ECA - Ant and post ethmoidal → ophthalmic. - Ophthalmic → ICA - Spheno palatine → maxillary. -
Peripheral Nervous System Peripheral Nervous System Comprises: A) Cranial Nerves: 12 in Number, Arise Mainly from Brain Stem
Peripheral nervous system Peripheral nervous system comprises: A) Cranial nerves: 12 in number, arise mainly from brain stem. B) Spinal nerves: arise from spinal cord, varies in number according to species. C) Autonomic nervous system : 1-Sympathetic (thoracolumbar) division. 2-Parasympathetic (craniosacral) division. Cranial nerves They are 12 pairs General features: – They are known by roman numbers from (rostral to caudal) . – Some nerves take their names according to: * Function as olfactory and abducent * Distribution as facial and hypoglossal * Shape as trigeminal. - All cranial nerves have superficial origins on ventral aspect of brain except trochlear nerve originates from dorsal aspect of brain. -All cranial nerves except first one originate from brain stem. - Each nerve emerges from cranial cavity through a foramen. - All cranial nerves except vagus distributed generally in head region. - 3,7,9,10 cranial nerves carry parasympathetic fibers. - Classification of cranial nerves: - They are classified according to the function into: 1-Sensory nerves: 1,2,8. 2-Motor nerves: 3,4,6,11,12. 3-Mixed nerves: 5,7,9,10. I- Olfactory nerves Type: • Sensory for smell. • Represented by bundles of nerve fibers, not form trunk. Origin: • Central processes of olfactory cells of olfactory region of nasal cavity. Course: • They pass though cribriform plate to join olfactory bulb. Vomeronasal nerve: • It arises from vomeronasal organ ,passes through medial border of cribriform plate to terminate in olfactory bulb. II- Optic nerve Type: • Sensory for vision. Origin: • Central processes of ganglion cells of retina. Course: • Fibers converge toward optic papilla forming optic nerve, emerges from eyeball. • Then passes through optic foramen and decussates with its fellow to form optic chiasma. -

European Position Paper on the Anatomical Terminology of the Internal Nose and Paranasal Sinuses
ISSN: 03000729 INTERN AT IO N A L R H I N CONTENT O L O G I C Official Journal of the European and International Societies Position paper Lund VJ, Stammberger H, Fokkens WJ, Beale T, Bernal-Sprekelsen M, Eloy P, Georgalas C, Ger- S O C I E Y stenberger C, Hellings PW, Herman P, Hosemann WG, Jankowski R, Jones N, Jorissen M, Leunig T A, Onerci M, Rimmer J, Rombaux P, Simmen D, Tomazic PV, Tschabitscher M, Welge-Luessen A. European Position Paper on the Anatomical Terminology of the Internal Nose and Parana- VOLUME 50 | SUPPLEMENT 24 | MARCH 2014 sal Sinuses. Rhinology. 2014 Suppl. 24: 1-34. European Position Paper on the Anatomical Terminology of the Internal Nose and Paranasal Sinuses Lund VJ, Stammberger H, Fokkens WJ et al. 2014 Anatomical terminology cover JS.indd 1 27-02-14 23:03 European Position Paper on the Anatomical Terminology of the INTERN AT Internal Nose and Paranasal Sinuses IO N A L R H I N O L O G I C Official Journal of the European and International Rhinologic Societies S O C I E Y T Editor-in-Chief Address Prof V.J. Lund Journal Rhinology, c/o AMC, Mrs. J. Kosman / A2-234, PO Box 22 660, Prof W.J. Fokkens 1100 DD Amsterdam, the Netherlands. Tel: +31-20-566 4534 Associate Editor Fax: +31-20-566 9662 Prof P.W. Hellings E-mail: [email protected] Website: www.rhinologyjournal.com Managing Editor Dr. W.T.V. Germeraad Assistant Editor Dr. Ch. Georgalas Editorial Assistant (contact for manuscripts) Mrs J. -

Vidian Canal: Radiological Anatomy and Functional Correlations
Original article Vidian canal: radiological anatomy and functional correlations Bidarkotimath, S.1*, Viveka, S.2 and Udyavar, A.3 1MBBS, MD (Anatomy), Assistant Professor, Department of Anatomy, A. J. Institute of Medical Sciences, 575004, Manglore, Índia 2Post Graduate student, Department of Anatomy, A. J. Institute of Anatomy, 575004, Mangalore, Índia 3MBBS, MD (Anatomy), Professor and Head, Department of Anatomy, A. J. Institute of Medical Sciences, 575004, Manglore, Índia *E-mail: [email protected]; [email protected]; [email protected] Abstract The Vidian or pterygoid canal is the osseous tunnel for both the Vidian artery and nerve. The aim of the current study is to delineate the Vidian canal in CT scans of head and to establish the landmarks for swift and easy localization of the canal. It is also intended to focus on the functional correlations of the structures present in the canal with its anatomy. The Vidian canal is studied in 200 adult CT scans and its measurements including its internal diameter, distances of the canal from the vomerine crest, from superior & inferior sphenoid sinus wall and from the foramen rotendum are noted. It is observed that canal starts from the medial end of petrous part of internal carotid or up to 8 mm from its medial end, mean distance being 6 mm and runs medial to lateral from foramen lacerum in 1/3rd of cases. Canal with suspended by bony stalk into sphenoid sinus noted in 22%, canal embedded in the body of sphenoid noted in 11% and rest had canals in normal expected position. In 5 instances the canal was found to be divided into two identifiable canals. -
The Pterygopalatine Fossa and Its Connections
Where is it? • Deep to the infratemporal fossa • Anterior to the pterygoid process • Behind the posterior wall of the maxillary sinus (perpendicular plate of the palatine bone) Pterygopalatine fossa and its connections David R. DeLone, MD ©2017 MFMER | slide-1 ©2017 MFMER | slide-2 What’s in it? What’s in it? ©2017 MFMER | slide-3 ©2017 MFMER | slide-4 Autonomic function What goes in and out of it? • Posterior • Parasympathetic • Foramen rotundum • Parasympathetic preganglionic fibers coming along Vidian nerve (GSPN) • Vidian canal synapse at pterygopalatine ganglion. • Palatovaginal (pharyngeal) canal • Postganglionic fibers pass along the pterygopalatine nerves, to maxillary nerve, zygomatic nerve (inferior orbital fissure), and then along • Vomerovaginal (basipharyngeal) canal zygomaticotemporal nerve. The fibers then communicate with the lacrimal • Medial branch of the ophthalmic nerve. • Sphenopalatine foramen • Supplies the lacrimal gland and mucosa of nasopharynx and nasal cavity. • Lateral • Sympathetic • Pterygomaxillary fissure • Postganglionic fibers arising from superior cervical ganglion pass along the carotid plexus, become deep petrosal nerve, which joins the GSPN to form • Anterosuperior the Vidian nerve. • Inferior orbital fissure • Sympathetic fibers pass through pterygopalatine ganglion without synapse and then distribute analogously to the parasympathetic fibers. • Inferior • Greater and lesser palatine foramina ©2017 MFMER | slide-5 ©2017 MFMER | slide-6 What goes in and out of it? • Posterior • Foramen rotundum (Meckel’s cave) • Maxillary nerve (V2) • Artery of foramen rotundum • Emissary veins • Vidian canal (foramen lacerum) • Vidian nerve and Vidian artery • Greater superficial petrosal nerve • Deep petrosal nerve • Palatovaginal (pharyngeal) canal • Pterygovaginal artery • Pharyngeal nerve to Eustachian tube • Vomerovaginal (basipharyngeal) canal • Branch of the sphenopalatine artery Vidus Vidius Tubbs Neurosurgery 2006 (Guido Guidi) Rumboldt AJR 2006 circa 1547. -

Skull – Communication
Multimedial Unit of Dept. of Anatomy JU Anterior cranial fossa The floor: ● Cribriform plate of ethmoid ● Orbital parts of frontal ● Lesser wings of sphenoid ● Body of sphenoid – anterior to prechiasmatic sulcus Contents: ● Frontal lobes of brain ● Olfactory bulbs ● Olfactory tracts ● Anterior meningeal vessels (from anterior ethmoid) Communication: 1. Through cribriform plate of ethmoid with the nasal cavity Contents: ● Olfactory fila ● Anterior athmoidal vessels and nerves 2. Through foramen cecum with the nasal cavity ● Extension of dura mater ● Small vein connecting veins of nasal cavity and superior sagittal sinus Middle cranial fossa It consists of the central (body of sphenoid) and two lateral parts. Lateral parts consist of: ● Greater wing of sphenoid ● Squamous parts of temporal ● Anterior surfaces of pyramids of temporal The border between the anterior and middle cranial fossa ● Posterior margins of lesser wings of sphenoid ● Sphenoidal limbus The border between the middle and posterior cranial fossae: ● Superior margins of the petrous parts of temporal ● Dorsum sellae Contents: Central part ● Interbrain ● Intercavernous sinuses Lateral part ● Temporal lobes of brain ● Cavernous sinus ● Cranial nerves II – VI ● Internal carotid arteries ● Middle meningeal vessels ● Greater and lesser petrosal nerves Cavernous sinus Contents: ● Internal carotid artery ● Cavernous plexus ● Abducens nerve ● Lateral wall of cavernous sinus contains: ● Oculomotor nerve ● Trochlear nerve ● Ophthalmic nerve ● Maxillary nerve Communication of middle cranial fossa: 1. Through superior orbital fissure with the orbit Contents: ● Oculomotor nerve ● Trochlear nerve ● Ophthalmic nerve ● Abducens nerve ● Sympathetic postganglionic axons of the cavernous plexus ● Superior ophthalmic vein ● Superior branch of the inferior ophthalmic vein ● Ramus of middle meningeal artery 2. Optic canal – with orbit ● Optic nerve ● Ophthalmic artery 3.