Lead Poisoning
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Erythrocytes: Overview, Morphology, Quantity by AH Rebar Et
In: A Guide to Hematology in Dogs and Cats, Rebar A.H., MacWilliams P.S., Feldman B.F., Metzger F.L., Pollock R.V.H. and Roche J. (Eds.). Publisher: Teton NewMedia, Jackson WY (www.veterinarywire.com). Internet Publisher: International Veterinary Information Service, Ithaca NY (www.ivis.org), 8-Feb-2005; A3304.0205 Erythrocytes: Overview, Morphology, Quantity A.H. Rebar1, P.S. MacWilliams2, B.F. Feldman 3, F.L. Metzger 4, R.V.H. Pollock 5 and J. Roche 6 1Dept of Veterinary Pathobiology, School of Veterinary Medicine, Purdue University, IN,USA. 2Dept of Pathobiological Sciences, School of Veterinary Medicine, University of Wisconsin, WI, USA. 3Dept of Biomedical Sciences & Pathobiology, VA-MD - Regional College of Veterinary Medicine, Virginia Tech, VA, USA. 4Metzger Animal Hospital,State College,PA, USA. 5Fort Hill Company, Montchanin, DE, USA. 6 Hematology Systems, IDEXX Laboratories, Westbrook, ME, USA. Overview Production Red blood cells (RBC) are produced in the bone marrow. Numbers of circulating RBCs are affected by changes in plasma volume, rate of RBC destruction or loss, splenic contraction, erythropoietin (EPO) secretion, and the rate of bone marrow production. A normal PCV is maintained by an endocrine loop that involves generation and release of erythropoietin (EPO) from the kidney in response to renal hypoxia. Erythropoietin stimulates platelet production as well as red cell production. However, erythropoietin does not stimulate white blood cell (WBC) production. Erythropoiesis and RBC numbers are also affected by hormones from the adrenal cortex, thyroid, ovary, testis, and anterior pituitary. Destruction Red cells have a finite circulating lifespan. In dogs, the average normal red cell circulates approximately 100 days. -

The Genetics of Metabolic Disorders* D
Postgrad Med J: first published as 10.1136/pgmj.48.558.207 on 1 April 1972. Downloaded from Postgraduate Medical Journal (April 1972) 48, 207-2,11. The genetics of metabolic disorders* D. A. HOPKINSON M.A., M.D. MRC Human Biochemical Genetics Unit, Galton Laboratory, University College London THE first work on the genetics of metabolic disorders tion (Harris, 1970) more than sixty such disorders in Man was of course carried out by that great were listed and further examples are regularly being pioneer Sir Archibald Garrod at the turn of the reported. present century (Garrod, 1902). From a study of the It is interesting to note that there is no one hereditary background of a small series of patients particular aspect of metabolism which seems with a rare disorder, alcaptonuria, he discovered the peculiarly susceptible to genetic variation of this first example of recessive Mendelian inheritance in kind. Nor is there any indication that enzymes Man and with remarkable foresight he suggested catalysing particular types of reaction or utilizing that this unusual condition was an example of specialized cofactors are more prone. On the one 'chemical individuality'-an inborn alteration in the hand we have disorders like acatalasia in which metabolism of tyrosine. He also suggested that other there is a deficiency of catalase, the enzyme res- conditions such as albinism and cystinuria could be ponsible for the simple reaction splitting hydrogen manifestations of inborn changes or defects in meta- peroxide to water and oxygen, and on the other hand bolism. He called them 'freaks' or 'sports' of meta- we have the glycogen diseases in which there are bolism. -

Download Download
Acta Biomed 2021; Vol. 92, N. 1: e2021169 DOI: 10.23750/abm.v92i1.11345 © Mattioli 1885 Update of adolescent medicine (Editor: Vincenzo De Sanctis) Rare anemias in adolescents Joan-Lluis Vives Corrons, Elena Krishnevskaya Red Blood Cell Pathology and Hematopoietic Disorders (Rare Anaemias Unit) Institute for Leukaemia Research Josep Car- reras (IJC). Badalona (Barcelona) Abstract. Anemia can be the consequence of a single disease or an expression of external factors mainly nutritional deficiencies. Genetic issues are important in the primary care of adolescents because a genetic diagnosis may not be made until adolescence, when the teenager presents with the first signs or symptoms of the condition. This situation is relatively frequent for rare anemias (RA) an important, and relatively heteroge- neous group of rare diseases (RD) where anaemia is the first and most relevant clinical manifestation. RA are characterised by their low prevalence (< 5 cases per 10,000 individuals), and, in some cases, by their complex mechanism. For these reasons, RA are little known, even among health professionals, and patients tend to re- main undiagnosed or misdiagnosed for long periods of time, making impossible to know the prognosis of the disease, or to carry out genetic counselling for future pregnancies. Since this situation is an important cause of anxiety for both adolescent patients and their families, the physician’s knowledge of the natural history of a genetic disease will be the key factor for the anticipatory guidance for diagnosis and clinical follow-up. RA can be due to three primary causes: 1. Bone marrow erythropoietic defects, 2. Excessive destruction of mature red blood cells (hemolysis), and 3. -

Haematological Abnormalities in Mitochondrial Disorders
Singapore Med J 2015; 56(7): 412-419 Original Article doi: 10.11622/smedj.2015112 Haematological abnormalities in mitochondrial disorders Josef Finsterer1, MD, PhD, Marlies Frank2, MD INTRODUCTION This study aimed to assess the kind of haematological abnormalities that are present in patients with mitochondrial disorders (MIDs) and the frequency of their occurrence. METHODS The blood cell counts of a cohort of patients with syndromic and non-syndromic MIDs were retrospectively reviewed. MIDs were classified as ‘definite’, ‘probable’ or ‘possible’ according to clinical presentation, instrumental findings, immunohistological findings on muscle biopsy, biochemical abnormalities of the respiratory chain and/or the results of genetic studies. Patients who had medical conditions other than MID that account for the haematological abnormalities were excluded. RESULTS A total of 46 patients (‘definite’ = 5; ‘probable’ = 9; ‘possible’ = 32) had haematological abnormalities attributable to MIDs. The most frequent haematological abnormality in patients with MIDs was anaemia. 27 patients had anaemia as their sole haematological problem. Anaemia was associated with thrombopenia (n = 4), thrombocytosis (n = 2), leucopenia (n = 2), and eosinophilia (n = 1). Anaemia was hypochromic and normocytic in 27 patients, hypochromic and microcytic in six patients, hyperchromic and macrocytic in two patients, and normochromic and microcytic in one patient. Among the 46 patients with a mitochondrial haematological abnormality, 78.3% had anaemia, 13.0% had thrombopenia, 8.7% had leucopenia and 8.7% had eosinophilia, alone or in combination with other haematological abnormalities. CONCLUSION MID should be considered if a patient’s abnormal blood cell counts (particularly those associated with anaemia, thrombopenia, leucopenia or eosinophilia) cannot be explained by established causes. -
Complete Blood Count in Primary Care
Complete Blood Count in Primary Care bpac nz better medicine Editorial Team bpacnz Tony Fraser 10 George Street Professor Murray Tilyard PO Box 6032, Dunedin Clinical Advisory Group phone 03 477 5418 Dr Dave Colquhoun Michele Cray free fax 0800 bpac nz Dr Rosemary Ikram www.bpac.org.nz Dr Peter Jensen Dr Cam Kyle Dr Chris Leathart Dr Lynn McBain Associate Professor Jim Reid Dr David Reith Professor Murray Tilyard Programme Development Team Noni Allison Rachael Clarke Rebecca Didham Terry Ehau Peter Ellison Dr Malcolm Kendall-Smith Dr Anne Marie Tangney Dr Trevor Walker Dr Sharyn Willis Dave Woods Report Development Team Justine Broadley Todd Gillies Lana Johnson Web Gordon Smith Design Michael Crawford Management and Administration Kaye Baldwin Tony Fraser Kyla Letman Professor Murray Tilyard Distribution Zane Lindon Lyn Thomlinson Colleen Witchall All information is intended for use by competent health care professionals and should be utilised in conjunction with © May 2008 pertinent clinical data. Contents Key points/purpose 2 Introduction 2 Background ▪ Haematopoiesis - Cell development 3 ▪ Limitations of reference ranges for the CBC 4 ▪ Borderline abnormal results must be interpreted in clinical context 4 ▪ History and clinical examination 4 White Cells ▪ Neutrophils 5 ▪ Lymphocytes 9 ▪ Monocytes 11 ▪ Basophils 12 ▪ Eosinophils 12 ▪ Platelets 13 Haemoglobin and red cell indices ▪ Low haemoglobin 15 ▪ Microcytic anaemia 15 ▪ Normocytic anaemia 16 ▪ Macrocytic anaemia 17 ▪ High haemoglobin 17 ▪ Other red cell indices 18 Summary Table 19 Glossary 20 This resource is a consensus document, developed with haematology and general practice input. We would like to thank: Dr Liam Fernyhough, Haematologist, Canterbury Health Laboratories Dr Chris Leathart, GP, Christchurch Dr Edward Theakston, Haematologist, Diagnostic Medlab Ltd We would like to acknowledge their advice, expertise and valuable feedback on this document. -

Red Blood Cell Morphology in Patients with Β-Thalassemia Minor
J Lab Med 2017; 41(1): 49–52 Short Communication Carolin Körber, Albert Wölfler, Manfred Neubauer and Christoph Robier* Red blood cell morphology in patients with β-thalassemia minor DOI 10.1515/labmed-2016-0052 Keywords: β-thalassemia minor; erythrocytes; red blood Received July 11, 2016; accepted October 20, 2016; previously published cells; red blood cell morphology. online December 10, 2016 Abstract In β-thalassemias, the examination of a peripheral blood (PB) smear may provide relevant clues to initial diagnosis. Background: A systematic analysis of the occurrence of Complete laboratory investigation consists of the determina- red blood cell (RBC) abnormalities in β-thalassemia minor tion of the complete blood count, assessment of red blood has not been performed to date. This study aimed to iden- cell (RBC) morphology, high performance liquid chroma- tify and quantify the frequency of RBC abnormalities in tography (HPLC), hemoglobin electrophoresis and, where patients with β-thalassemia minor. necessary, DNA analysis [1]. Especially in the clinically Methods: We examined blood smears of 33 patients with severe forms referred to as β-thalassemia major and interme- β-thalassemia minor by light microscopy for the occur- dia, RBC abnormalities are often markedly apparent [2]. In rence of 15 defined RBC abnormalities. In the case of posi- β-thalassemia minor, also called β-thalassemia trait, the car- tivity, the abnormal cells/20 high power fields (HPF) at riers are usually clinically asymptomatic, showing persistent 1000-fold magnification were counted. microcytosis and hypochromia or mild microcytic anemia [1, Results: Anisocytosis, poikilocytosis and target cells 3]. The PB smear may show microcytosis, hypochromia and, (median 42/20 HPF) were observed in all, and ovalocytes infrequently, poikilocytosis [2]. -

Chapter 03- Diseases of the Blood and Certain Disorders Involving The
Chapter 3 Diseases of the blood and blood-forming organs and certain disorders involving the immune mechanism (D50- D89) Excludes2: autoimmune disease (systemic) NOS (M35.9) certain conditions originating in the perinatal period (P00-P96) complications of pregnancy, childbirth and the puerperium (O00-O9A) congenital malformations, deformations and chromosomal abnormalities (Q00-Q99) endocrine, nutritional and metabolic diseases (E00-E88) human immunodeficiency virus [HIV] disease (B20) injury, poisoning and certain other consequences of external causes (S00-T88) neoplasms (C00-D49) symptoms, signs and abnormal clinical and laboratory findings, not elsewhere classified (R00-R94) This chapter contains the following blocks: D50-D53 Nutritional anemias D55-D59 Hemolytic anemias D60-D64 Aplastic and other anemias and other bone marrow failure syndromes D65-D69 Coagulation defects, purpura and other hemorrhagic conditions D70-D77 Other disorders of blood and blood-forming organs D78 Intraoperative and postprocedural complications of the spleen D80-D89 Certain disorders involving the immune mechanism Nutritional anemias (D50-D53) D50 Iron deficiency anemia Includes: asiderotic anemia hypochromic anemia D50.0 Iron deficiency anemia secondary to blood loss (chronic) Posthemorrhagic anemia (chronic) Excludes1: acute posthemorrhagic anemia (D62) congenital anemia from fetal blood loss (P61.3) D50.1 Sideropenic dysphagia Kelly-Paterson syndrome Plummer-Vinson syndrome D50.8 Other iron deficiency anemias Iron deficiency anemia due to inadequate dietary -
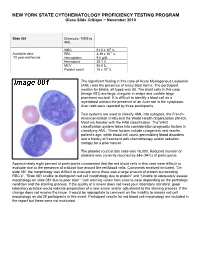
10 11 Cyto Slides 81-85
NEW YORK STATE CYTOHEMATOLOGY PROFICIENCY TESTING PROGRAM Glass Slide Critique ~ November 2010 Slide 081 Diagnosis: MDS to AML 9 WBC 51.0 x 10 /L 12 Available data: RBC 3.39 x 10 /L 72 year-old female Hemoglobin 9.6 g/dL Hematocrit 29.1 % MCV 86.0 fL Platelet count 16 x 109 /L The significant finding in this case of Acute Myelogenous Leukemia (AML) was the presence of many blast forms. The participant median for blasts, all types was 88. The blast cells in this case (Image 081) are large, irregular in shape and contain large prominent nucleoli. It is difficult to identify a blast cell as a myeloblast without the presence of an Auer rod in the cytoplasm. Auer rods were reported by three participants. Two systems are used to classify AML into subtypes, the French- American-British (FAB) and the World Health Organization (WHO). Most are familiar with the FAB classification. The WHO classification system takes into consideration prognostic factors in classifying AML. These factors include cytogenetic test results, patient’s age, white blood cell count, pre-existing blood disorders and a history of treatment with chemotherapy and/or radiation therapy for a prior cancer. The platelet count in this case was 16,000. Reduced number of platelets was correctly reported by 346 (94%) of participants. Approximately eight percent of participants commented that the red blood cells in this case were difficult to evaluate due to the presence of a bluish hue around the red blood cells. Comments received included, “On slide 081 the morphology was difficult to evaluate since there was a large amount of protein surrounding RBC’s”, “Slide 081 unable to distinguish red cell morphology due to protein” and “Unable to adequately assess morphology on slide 081 due to poor stain”. -

Clinical Consequences of Enzyme Deficiencies in the Erythrocyte
ANNALS OF CLINICAL AND LABORATORY SCIENCE, Vol. 10, No. 5 Copyright © 1980, Institute for Clinical Science, Inc. Clinical Consequences of Enzyme Deficiencies in the Erythrocyte MICHAEL L. NETZLOFF, M.D. Department of Pediatrics and Human Development, Michigan State University College of Human Medicine, East Lansing, Ml 48824 ABSTRACT The anueleate mature erythrocyte also lacks ribosomes and mitochondria and thus cannot synthesize enzymes or derive energy from the Krebs citric acid cycle. Nevertheless, the red blood cell is metabolically active and contains numerous residual enzymes and their products which are essential for its survival and normal functioning. Enzyme deficiencies in the Embden-Myerhoff glycolytic pathway can result in nonspherocytic hemoly tic anemia (NSHA), and some are also associated with neuromuscular or neurologic disorders. Glucose-6-phosphate dehydrogenase deficiency in the hexose monophosphate shunt also results in hemolytic anemia, espe cially following exposure to various drugs. Defects in glutathione synthesis and pyrimidine 5'-nucleotidase deficiency also cause NSHA, as does in creased adenosine deaminase activity. Glutathione synthetase deficiency which is not limited to the red cell also presents as oxoprolinuria with neurologic signs. All red cell enzyme defects appear as single gene errors, in most cases recessive in inheritance, either autosomal or X-linked. Clinical Consequences of Enzyme tains over 40 different enzymes, many of Deficiencies in the Erythrocyte which are essential for its survival or nor mal functioning. The purpose of this re Prior to maturity, the nucleated red port is to review some inherited enzyme blood cell (rbc) can perform a variety of deficiencies of the erythrocyte which metabolic functions, including active en compromise its viability in the circulation zyme synthesis. -

Hemolytic Anemia with Impaired Hexokinase Activity
Hemolytic anemia with impaired hexokinase activity Alan S. Keitt J Clin Invest. 1969;48(11):1997-2007. https://doi.org/10.1172/JCI106165. Research Article Analyses of key glycolytic intermediates in freshly drawn red cells from six related individuals suggest that decreased hexokinase activity underlies the hemolytic process in the two members with overt hemolysis. Low red cell glucose 6- phosphate (G6P) was observed not only in the anemic patients but in the presumptive heterozygotes as well and served as a useful marker for the presence of the trait. Hexokinase activity was labile in distilled water hemolysates but was only slightly low when protected by glucose, mercaptoethanol, and ethylenediaminetetraacetate (EDTA). Normal red cell hexokinase was demonstrated to be dependent on glucose for maintenance of activity after heating to 45°C. The cells of the proposita are unable to utilize glucose efficiently at glucose concentrations lower than 0.2 mmole/liter whereas normal cells maintain linear glucose consumption to at least 0.05 mM glucose. These qualitative abnormalities could result from the presence of a mutant hexokinase with an abnormally reactive sulfhydryl group and altered substrate affinity in the red cells of this kindred. Find the latest version: https://jci.me/106165/pdf Hemolytic Anemia with Impaired Hexokinase Activity ALAN S. KErrr From the Department of Medicine, University of Florida College of Medicine, Gainesville, Florida 32601 A B S T R A C T Analyses of key glycolytic intermediates cells of a young girl with moderately severe chronic in freshly drawn red cells from six related individuals anemia. Although hexokinase activity was only slightly suggest that decreased hexokinase activity underlies the below the range of normal, a comparison between the hemolytic process in the two members with overt he- activity of red cell hexokinase in the affected proposita molysis. -

Research Article
z Available online at http://www.journalcra.com INTERNATIONAL JOURNAL OF CURRENT RESEARCH International Journal of Current Research Vol. 9, Issue, 12, pp.62497-62502, December, 2017 ISSN: 0975-833X RESEARCH ARTICLE INCLUSION BODIES - A REVIEW 1Dr. Mrunalini Mahesh Dadpe, 2, *Dr. Sourab Kumar, 3Dr. Mahesh Dadpe, 4Dr. Payoshnee Bhalinge Jadhav, 5Dr. Abhishek Jadhav and 6Dr. Shilpi Suman 1, 4Department of Oral Pathology, M A Rangoonwala Dental College, Aazam Campus, Camp, Pune 411001, India 2, 5, 6Department of oral Pathology, D.Y. Patil School of Dentistry, Nerul, Navi Mumbai 400706, India 3MIDSR Dental College, Vishwanathpuram, Amba Jogai Road, Latur 413531, India ARTICLE INFO ABSTRACT Article History: The word inclusion means incorporation. Inclusion bodies are nuclear or cytoplasmic aggregates of Received 16th September, 2017 stainable substances which are usually ‘proteins’. They typically represent sites of viral multiplication Received in revised form in bacteria or in a eukaryotic cell and usually consist of viral capsid proteins. Inclusion bodies are 17th October, 2017 typically identified within a cell by their different appearance. Accepted 25th November, 2017 Published online 27th December, 2017 Key words: Proteins, Viral, Types, Structures Copyright © 2017, Mrunalini Mahesh Dadpe et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Citation: Dr. Mrunalini Mahesh Dadpe, Dr. Sourab Kumar, Dr. Mahesh Dadpe, Dr. Payoshnee Bhalinge Jadhav, Dr. Abhishek Jadhav and Dr. Shilpi Suman, 2017. “Inclusion Bodies - A Review”, International Journal of Current Research, 9, (12), 62497-62502. Classification INTRODUCTION Viral inclusion bodies Inclusion bodies (IB) can also be hallmarks of genetic diseases, as in the case of Neuronal Inclusion bodies in neural disorders, A. -
Blood Lab First Week
Blood Lab First Week This is a self propelled powerpoint study atlas of blood cells encountered in examination of the peripheral blood smear. It is a requirement of this course that you begin review of these slides with the first day of class in order to familiarize yourself with terms used in describing peripheral blood morphology. Laboratory final exam questions are derived directly from these slides. The content of these slides may duplicate slides (1-37) listed on the Hematopathology Website. You must familiarize yourself with both sets of slides in order to have the best learning opportunity. Before Beginning This Slide Review • Please read the Laboratory information PDF under Laboratory Resources file to learn about – Preparation of a blood smear – How to select an area of the blood smear for review of morphology and how to perform a white blood cell differential – Platelet estimation – RBC morphology descriptors • Anisocytosis • Poikilocytosis • Hypochromia • Polychromatophilia Normal blood smear. Red blood cells display normal orange pink cytoplasm with central pallor 1/3-1/2 the diameter of the red cell. There is mild variation in size (anisocytosis) but no real variation in shape (poikilocytosis). To the left is a lymphocyte. To the right is a typical neutrophil with the usual 3 segmentations of the nucleus. Med. Utah pathology Normal blood: thin area Ref 2 Normal peripheral blood smear. This field is good for exam of cell morphology, although there are a few minor areas of overlap of red cells. Note that most cells are well dispersed and the normal red blood cell central pallor is noted throughout the smear.