Plant Systematics
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Tasmannia Lanceolata
ASPECTS OF LEAF AND EXTRACT PRODUCTION from Tasmannia lanceolata by Chris Read, B. Agr.Sc. Tas. Submitted in fulfillment of the requirements for the Degree of Doctor of Philosophy University of Tasmania, Hobart December 1995 ' s~, ... ~~ \ ·'(11 a_C\14 \t\J. \I ' This thesis contains no material which has been accepted for the award of any other degree or diploma in any University, and to the best of my knowledge, contains no copy or paraphrase of material previously written or published by any other person except where due reference is given in the text. University of Tasmania HOBART March 1996 This thesis may be made available for loan and limited copying in accordance with the Copyright Act 1968 University of Tasmania HOBART March 1996 Abstract This thesis examines several aspects of the preparation, extraction and analysis of solvent soluble compounds from leaf material of Tasmannia lanceolata and reports a preliminary survey of extracts of some members of the natural population of the species in Tasmania. A major constituent of these extracts, polygodial, was shown to be stored within specialised idioblastic structures scattered throughout the mesophyll, and characterised by distinctive size and shape, and a thickened wall. The contents of these cells were sampled directly, analysed and compared with the composition of extracts derived from ground, dry whole leaf. This result was supported by spectroscopic analysis of undisturbed oil cells in whole leaf tissue. In a two year field trial, the progressive accumulation of a number of leaf extract constituents (linalool, cubebene, caryophyllene, germacrene D, bicyclogermacrene, cadina-1,4 - diene, aristolone and polygodial) during the growth flush was followed by a slow decline during the subsequent dormant season. -

And According to Bailey & (1943)
BLUMEA 24 (1978) 521—525 The Winteraceae pf the Old World. III. Notes on the ovary of Takhtajania W. Vink Rijksherbarium, Leiden, Netherlands INTRODUCTION Very recently Baranova & Leroy (Leroy, 1978) published a new genus Takhta- jania to accomodate the aberrant Bubbia perrieri Capuron. The outstanding characters of this genus are the anomocytic stomatal apparatus (Baranova, 1972; Bongers, 1973) and the unilocular bicarpellate ovary (Leroy, 1977, 1978). work the Winteraceae, also studied the of Bub- During my on I single specimen bia perrieri in existence. The late Capuron told me that he had tried to collect additional specimens but that he had not succeeded in doing so; according to him the type locality was completely deforested. In view of the scarcity of the material it was considered relevant to publish some additional notes without delay. MATERIAL AND METHODS I studied three ovaries of the type specimen (Perrier de la Bdthie 10158). One was cut longitudinally, slightly outside the plane of symmetry; another was cut and its half These drawn transversally upper again longitudinally. parts were (figs. 2 and 3) and then cleared according to Bailey & Nast (1943) (figs 5 and third boiled and then sectioned 6). The ovary was serially (fig. 4). OBSERVATIONS One of the ovaries was taken from a flowerbud of which the closed petals were 57 mm high. The diagram of this flowerbud is drawn in fig. 1. The 2 slightly ruptured calyx showed three putative original apices ± alternating with the three bracteoles at the base of the pedicel. The petals are all free. The ovary is its and the flattened, has a longitudinal groove on narrow sides, highest sta- mens are inserted adjoining its broadest sides. -

<I>Winteraceae</I>
Blumea 59, 2014: 155–162 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE http://dx.doi.org/10.3767/000651914X686022 The Winteraceae of the Old World. VII. Zygogynum in the Solomon Islands (incl. Bougainville) W. Vink1 Key words Abstract A review of the character distribution in the Zygogynum representatives from the Solomon Islands (and Bougainville) shows a rather wide variability in ranges of numbers and sizes, often with distinct differences between Solomon Islands islands or altitudinal ranges. However, the morphology is rather uniform. The four species recognised by A.C. Smith Winteraceae had to be synonymised, but a new taxon deserved recognition. The name Bubbia is explained. Zygogynum Published on 19 December 2014 INTRODUCTION CHARACTERS AND THEIR VARIABILITY The first Zygogynum species described from the Solomon Is- Leaves lands is Bubbia haplopus B.L.Burtt (1936). In his monumental Except for size, the leaves are very uniform with the blade works on the Papuasian plants, Smith (1942) transferred this chartaceous, the midrib impressed above and prominent below. species from Bubbia to the other Van Tieghem genus, Belliolum, On the lower side of the blade the grey to white alveolar mate- and described three more species from this area. The materials rial covers only the stomata or unites these into small strings available to Smith were, however, very incomplete, and several or clusters; the stomata are also present over the secondary characters could not be established; next to leaf size, minor nerves, or less dense to lacking over the thickest parts of the differences had to be used to define these taxa. -

Floristic Quality Assessment Report
FLORISTIC QUALITY ASSESSMENT IN INDIANA: THE CONCEPT, USE, AND DEVELOPMENT OF COEFFICIENTS OF CONSERVATISM Tulip poplar (Liriodendron tulipifera) the State tree of Indiana June 2004 Final Report for ARN A305-4-53 EPA Wetland Program Development Grant CD975586-01 Prepared by: Paul E. Rothrock, Ph.D. Taylor University Upland, IN 46989-1001 Introduction Since the early nineteenth century the Indiana landscape has undergone a massive transformation (Jackson 1997). In the pre-settlement period, Indiana was an almost unbroken blanket of forests, prairies, and wetlands. Much of the land was cleared, plowed, or drained for lumber, the raising of crops, and a range of urban and industrial activities. Indiana’s native biota is now restricted to relatively small and often isolated tracts across the State. This fragmentation and reduction of the State’s biological diversity has challenged Hoosiers to look carefully at how to monitor further changes within our remnant natural communities and how to effectively conserve and even restore many of these valuable places within our State. To meet this monitoring, conservation, and restoration challenge, one needs to develop a variety of appropriate analytical tools. Ideally these techniques should be simple to learn and apply, give consistent results between different observers, and be repeatable. Floristic Assessment, which includes metrics such as the Floristic Quality Index (FQI) and Mean C values, has gained wide acceptance among environmental scientists and decision-makers, land stewards, and restoration ecologists in Indiana’s neighboring states and regions: Illinois (Taft et al. 1997), Michigan (Herman et al. 1996), Missouri (Ladd 1996), and Wisconsin (Bernthal 2003) as well as northern Ohio (Andreas 1993) and southern Ontario (Oldham et al. -

Phytochemistry and Biological Properties of Drimys Winteri JR Et G. Forster Var Chilensis (DC) A
BOLETIN LATINOAMERICANO Y DEL CARIBE DE PLANTAS MEDICINALES Y AROMÁTICAS © / ISSN 0717 7917 / www.blacpma.ms-editions.cl Revisión / Review Phytochemistry and biological properties of Drimys winteri JR et G. Forster var chilensis (DC) A. [Fitoquímica y propiedades biológicas de Drimys winteri JR et G. Forster var chilensis (DC) A.] Orlando Muñoz1, Jorge Tapia-Merino2, Wolf Nevermann, & Aurelio San-Martín3 1Departamento de Quimica, Facultad de Ciencias, Universidad de Chile, Ñuñoa, Santiago, Chile 2Departamento de Ciencias Químicas y Biológicas, Facultad de Salud, Universidad Bernardo OHiggins, Santiago, Chile 3Departamento de Ciencias y Recursos Naturales, Facultad de Ciencias, Universidad de Magallanes, Punta Arenas, Chile Abstract: Drimys winteri JR et G. Forster var chilensis (DC) A. is a tree native to central and southern Chile. Also it found in part of Argentina. It is abundant in wet swampy localities from sea level to an altitude of 1700 m. This tree is sacred for the Mapuche culture; it is used in folk medicine in such as Reviewed by: inflammatory and painful processes. Phytochemical studies have demonstrated that this plant contains Arnaldo Bandoni Universidad de Buenos Aires mainly sesquiterpenes of the drimane type, flavonoids, essential oils, phytosterols and some lignans. Argentina These drimanes have attracted interest because of their antifeedant, plant growth regulation, cytotoxic, antimicrobial and insecticidal properties. The objective of this review is to establish clearly the Ali Parlar phytochemistry and biological activity of Drimys winteri JR et G. Forster var chilensis (DC) A. Articles Adiyaman University based on other varieties are not considered. Turkey Keywords: Drimys winteri; Sesquiterpenes; Essential oils; Lignans; Flavonoids; Biological properties. -

The Hoosier- Shawnee Ecological Assessment Area
United States Department of Agriculture The Hoosier- Forest Service Shawnee Ecological North Central Assessment Research Station General Frank R. Thompson, III, Editor Technical Report NC-244 Thompson, Frank R., III, ed 2004. The Hoosier-Shawnee Ecological Assessment. Gen. Tech. Rep. NC-244. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Research Station. 267 p. This report is a scientific assessment of the characteristic composition, structure, and processes of ecosystems in the southern one-third of Illinois and Indiana and a small part of western Kentucky. It includes chapters on ecological sections and soils, water resources, forest, plants and communities, aquatic animals, terrestrial animals, forest diseases and pests, and exotic animals. The information presented provides a context for land and resource management planning on the Hoosier and Shawnee National Forests. ––––––––––––––––––––––––––– Key Words: crayfish, current conditions, communities, exotics, fish, forests, Hoosier National Forest, mussels, plants, Shawnee National Forest, soils, water resources, wildlife. Cover photograph: Camel Rock in Garden of the Gods Recreation Area, with Shawnee Hills and Garden of the Gods Wilderness in the back- ground, Shawnee National Forest, Illinois. Contents Preface....................................................................................................................... II North Central Research Station USDA Forest Service Acknowledgments ................................................................................................... -
M23 Tree History Workbook Pub 2018
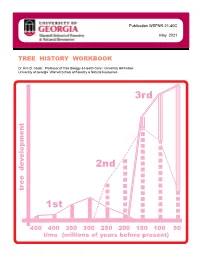
Publication WSFNR-21-40C May 2021 TREE HISTORY WORKBOOK Dr. Kim D. Coder, Professor of Tree Biology & Health Care / University Hill Fellow University of Georgia Warnell School of Forestry & Natural Resources 3rd 2nd tree development 1st 450 400 350 300 250 200 150 100 50 time (millions of years before present) This workbook is an educational product designed for helping people interested in trees to appreciate and understand how modern trees developed across long time periods. This product is a synthesis and integration of research and educational concepts regarding tree evolution and species inception. This educational workbook was designed for awareness building and foundation training of professional tree health care providers. At the time it was finished, this workbook contained educational materials and models concerning tree development thought by the author to provide the best means for considering the most basic understandings of time impacts on tree species creation, extinction, and tree biological and mechanical fundamentals. The University of Georgia, the Warnell School of Forestry & Natural Resources, and the author are not responsible for any errors, omissions, misinterpretations, or misapplications stemming from this educational product. The author assumed all users would have some basic tree biological, structural, and ecological background. This product was not designed, nor is suited, as a literature or research review. Please use the selected literature section at the back of this workbook to find scientific sources. This publication is copyrighted by the author. This educational product is only for noncommercial, nonprofit use and may not be copied or reproduced in whole or in part, by any means, in any format, or in any media including electronic forms, without explicit written permission of the author. -

Proceedings of the Indiana Academy of Science
List of Extirpated, Endangered, Threatened and Rare Vascular Plants in Indiana: An Update James R. Aldrich, John A. Bacone and Michael A. Homoya Indiana Department of Natural Resources Indianapolis, Indiana 46204 Introduction The status of Indiana's rarest vascular plants was last revised in 1981 by Bacone et al. (3). Since that publication much additional field work has been undertaken and accordingly, our knowledge of Indiana's rarest plant species has greatly increased. The results of some of this extensive field work have been discussed by Homoya (9) and Homoya and Abrell (10) for southern Indiana and by Aldrich et al. (1) for northern Indiana. Wilhelm's (17) discussion on the special vegetation of the Indiana Dunes Na- tional Lakeshore has also provided us with a much clearer understanding of the status of rare, threatened and endangered plants in northwest Indiana. Unfortunately, the number of species thought to be extinct in Indiana has more than tripled. Previous reports (2, 3) indicated that 26 species were extirpated in In- diana. The work that has been conducted to date leads us to believe that as many as 90 species may be extirpated. Without a doubt, the single factor most responsible for this extirpation has been and continues to be, the destruction of natural habitat. Compilation and Selection Criteria Lists of Bacone and Hedge (2), Bacone et al. (3), Barnes (4), Crankshaw (5) and Crovello (6) were consulted and provide the foundation for this report. New additions to this list include state records discovered since Deam (7) and a number of recommen- dations from field botanists. -

Recommended Native Pollinator-Friendly Plant List (Updated April 2021)
RECOMMENDED NATIVE POLLINATOR-FRIENDLY PLANT LIST (UPDATED APRIL 2021) Asheville GreenWorks is excited to share this updated native pollinator-friendly plant list for Asheville’s Bee City USA program! As the launchpad of the national Bee City USA program in 2012, we are gratified that throughout our community, individuals, organizations, and businesses are doing their part to reverse staggering global pollinator declines. Please check out our Pollinator Habitat Certification program at https://www.ashevillegreenworks.org/pollinator-garden-certification.html and our annual Pollination Celebration! during National Pollinator Week in June at https://www.ashevillegreenworks.org/pollination-celebration.html. WHY LANDSCAPE WITH POLLINATORS IN MIND? Asheville GreenWorks’ Bee City USA program encourages everyone to incorporate as many native plants into their landscapes and avoid insect-killing pesticides as much as possible. Here’s why. Over the millennia, hundreds of thousands of plant and animal pollinator species have perfected their pollination dances. Pollinating animals rely upon the carbohydrate-rich nectar and/or the protein-rich pollen supplied by flowers, and plants rely on pollinators to carry their pollen to other flowers to produce seeds and sustain their species. Nearly 90% of the world’s flowering plant species depend on pollinators to help them reproduce! Plants and pollinators form the foundation for our planet’s rich biodiversity generally. For example, 96% of terrestrial birds feed their young exclusively moth and butterfly caterpillars. ABOUT THIS NATIVE PLANT LIST An elite task force, listed at the end of this document, verified which plants were native to Western North Carolina and agreed this list should focus on plants’ value to pollinators as food--including nectar, pollen, and larval host plants for moth and butterfly caterpillars, as well as nesting habitat for bumble and other bees. -

Djvu Document
Cenomanian Angiosperm Leaf Megafossils, Dakota Formation, Rose Creek Locality, Jefferson County, Southeastern Nebraska By GARLAND R. UPCHURCH, JR. and DAVID L. DILCHER U.S. GEOLOGICAL SURVEY BULLETIN 1915 DEPARTMENT OF THE INTERIOR MANUEL LUJAN, JR., Secretary U.S. GEOLOGICAL SURVEY Dallas L. Peck, Director Any use of trade, product, or firm names in this publication is for descriptive purposes only and does not imply endorsement by the U.S. Government. UNITED STATES GOVERNMENT PRINTING OFFICE: 1990 For sale by the Books and Open-File Reports Section U.S. Geological Survey Federal Center Box 25425 Denver. CO 80225 Library of Congress Cataloging-in-Publication Data Upchurch, Garland R. Cenomanian angiosperm leaf megafossils, Dakota Formation, Rose Creek locality, Jefferson County, southeastern Nebraska / by Garland R. Upchurch, Jr., and David L. Dilcher. p. cm.-(U.S. Geological Survey bulletin; 1915) Includes bibliographical references. Supt. of Docs. no.: 1 19.3:1915. 1. Leaves, Fossil-Nebraska-Jefferson County. 2. Paleobotany-Cretaceous. 3. Paleobotany-Nebraska-Jefferson County. I. Dilcher, David L. II. Title. III. Series. QE75.B9 no. 1915 [QE983] 557.3 s-dc20 90-2855 [561'.2]CIP CONTENTS Abstract 1 Introduction 1 Acknowledgments 2 Materials and methods 2 Criteria for classification 3 Geological setting and description of fossil plant locality 4 Floristic composition 7 Evolutionary considerations 8 Ecological considerations 9 Key to leaf types at Rose Creek 10 Systematics 12 Magnoliales 12 Laurales 13 cf. Illiciales 30 Magnoliidae order -

Intercontinental Long-Distance Dispersal of Canellaceae from the New to the Old World Revealed by a Nuclear Single Copy Gene and Chloroplast Loci
Molecular Phylogenetics and Evolution 84 (2015) 205–219 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Intercontinental long-distance dispersal of Canellaceae from the New to the Old World revealed by a nuclear single copy gene and chloroplast loci Sebastian Müller a,1, Karsten Salomo a,1, Jackeline Salazar b, Julia Naumann a, M. Alejandra Jaramillo c, ⇑ Christoph Neinhuis a, Taylor S. Feild d,2, Stefan Wanke a, ,2 a Technische Universität Dresden, Institut für Botanik, Zellescher Weg 20b, 01062 Dresden, Germany b Escuela de Biología, Universidad Autónoma de Santo Domingo (UASD), C/Bartolomé Mitre, Santo Domingo, Dominican Republic c Centro de Investigación para el Manejo Ambiental y el Desarrollo, Cali, Colombia d Centre for Tropical Biodiversity and Climate Change, College of Marine and Environmental Science, Townsville 4810, Campus Townsville, Australia article info abstract Article history: Canellales, a clade consisting of Winteraceae and Canellaceae, represent the smallest order of magnoliid Received 10 July 2014 angiosperms. The clade shows a broad distribution throughout the Southern Hemisphere, across a diverse Revised 16 December 2014 range of dry to wet tropical forests. In contrast to their sister-group, Winteraceae, the phylogenetic rela- Accepted 17 December 2014 tions and biogeography within Canellaceae remain poorly studied. Here we present the phylogenetic Available online 9 January 2015 relationships of all currently recognized genera of Canellales with a special focus on the Old World Canellaceae using a combined dataset consisting of the chloroplast trnK-matK-trnK-psbA and the nuclear Keywords: single copy gene mag1 (Maigo 1). Within Canellaceae we found high statistical support for the mono- Canellales phyly of Warburgia and Cinnamosma. -

12939.Full.Pdf
Commentary Progress in understanding angiosperm history, success, and relationships: Darwin’s abominably ‘‘perplexing phenomenon’’ William L. Crepet* Department of Plant Biology, 228 Plant Science Building, Cornell University, Ithaca, NY 14853-4301 o Charles Darwin the suddenness of the Tangiosperm appearance and their rapid rise to dominance in the fossil record was both a ‘‘perplexing phenomenon’’ to ‘‘those who believe in extremely gradual evolution’’ and an ‘‘abominable mystery’’ (1). It has been 125 years since Darwin’s letter to Heer and for most of this time the investigation of the mystery has been in the domain of comparative morphology, traditional taxon- omy, and the fossil record, principally of Fig. 1. (a) A typical morphology-based phylogeny of existing seed plants (plus the extinct Bennettitales) pollen and leaves. These approaches have illustrating (in bold type) the anthophyte clade. (b) A composite phylogeny illustrating the realignment failed to illuminate the mystery that has now of Bennettitales and Gnetales based on ITS and new morphological data (21). grown to include major aspects of angio- sperm phylogeny, evolutionary success, and origin (2). But lately, exciting new methods and provide a precise framework within answer’’ (15), even though it conflicted and data are available that have enormous which to intelligently direct and ethically with his own previous analyses (16). potential to resolve this set of problems. evaluate the inevitable, if controversial, bio- Now a year later, in this issue of PNAS New kinds of fossils have been discovered engineering of plants for agricultural and Barkman et al. (17) report yet another but COMMENTARY from critical times (2–4); new algorithms are medicinal purposes.