Technical Paper 6
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ecological and Ethnobotanical Facet of 'Kelapa Hutan' (Pandanus Spp
OPEN ACCESS International Journal of Botany ISSN 1811-9700 DOI: 10.3923/ijb.2017.103.114 Research Article Ecological and Ethnobotanical Facet of ‘Kelapa Hutan’ (Pandanus Spp.) and Perspectives Towards its Existence and Benefit 1Krisma Lekitoo, 2Hans Fence Zakeus Peday, 2Novita Panambe and 2Reinardus Liborius Cabuy 1Manokwari Environmental and Forest Research Development Institute, 98314 Manokwari, West Papua Province, Indonesia 2Faculty of Forestry, University of Papua, Gunung Salju St., Amban, 98314 Manokwari, West Papua Province, Indonesia Abstract Background and Objective: Pandanus species are spread across the tropical New Guinea Forest and have long been extracted for food. In this study, the ecological and ethnobotanical aspects of Pandanus spp. were investigated. The objectives of the study were to highlight types of edible Pandanus spp. through their taxonomical characteristics, ecological distribution and fruit properties and to describe how traditional communities manage their existence and traditional values by way of ethnobotany and conservation. Methodology: To identify the potency and distribution of the edible Pandanus, continuous strip-sampling was applied to the sampling plots with an intensity of 5%. Temperature and humidity were measured directly under trees. The edible Pandanus specimens were identified by key experts and identification books. The thermogravimetric and Kjeldahl methods were implemented to identify nutrient contents. A semi-structural interview was implemented, which included questions on the management of Pandanus fruit and its social status among communities. Pearson’s correlation analysis was implemented to identify any relationship between temperature, humidity and fruit productivity. This analysis was performed using R statistical program. Results: Two edible Pandanus species were identified based on each characteristic: Pandanus brosimos Merr. -
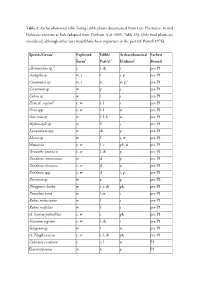
Table 2. Archaeobotanical Table Listing Edible Plants Documented from Late Pleistocene to Mid- Holocene Contexts at Kuk (Adapted from Denham Et Al
Table 2. Archaeobotanical table listing edible plants documented from Late Pleistocene to mid- Holocene contexts at Kuk (adapted from Denham et al. 2003: Table S3). Only food plants are considered, although other uses would have been important in the past (cf. Powell 1976). Species/Genus1 Exploited Edible Archaeobotanical Earliest Form2 Part(s)3 Evidence4 Record Abelmoschus sp. 5 c l, sh s pre-P1 Acalypha sp. w, t l s, p pre-P1 Castanopsis sp. w, t n w, p? pre-P1 Cerastium sp. w ps pre-P1 Coleus sp. w ls pre-P1 Ficus cf. copiosa5 c, w f, l s pre-P1 Ficus spp. c, w l, f w pre-P1 Garcinia sp. w f, l, b w pre-P1 Hydrocotyle sp. w l? s pre-P1 Lycopodium spp. w sh p pre-P1 Maesa sp. w f s, w pre-P1 Musaceae c, w f, c ph, st pre-P1 Oenanthe javanica c, w l, sh p pre-P1 Pandanus antaresensis w dp pre-P1 Pandanus brosimos c, w d p pre-P1 Pandanus spp. c, w d s, p pre-P1 Parsonsia sp. w np pre-P1 Phragmites karka w l, r, sh ph pre-P1 Pouzolzia hirta w l, st s pre-P1 Rubus moluccanus w fs pre-P1 Rubus rosifolius w fs pre-P1 cf. Setaria palmifolia c, w s ph pre-P1 Solanum nigrum c, w l, sh s pre-P1 Syzygium sp. w f w pre-P1 cf. Zingiberaceae c, w r, l, sh ph pre-P1 Colocasia esculenta c c, l st P1 Elaeocarpaceae w np P1 Species/Genus1 Exploited Edible Archaeobotanical Earliest Form2 Part(s)3 Evidence4 Record Ipomoea sp. -

Bioactive Components of Pandan's Fruits from Jayawijaya Mountains
IOSR Journal of Environmental Science, Toxicology and Food Technology (IOSR-JESTFT) e-ISSN: 2319-2402,p- ISSN: 2319-2399.Volume 8, Issue 8 Ver. I (Aug. 2014), PP 01-08 www.iosrjournals.org Bioactive Components of Pandan’s Fruits from Jayawijaya Mountains, Papua, Indonesia Been Kogoya1), Bambang Guritno2) Ariffin2) and Agus Suryanto 2) 1(Graduate School of Agriculture, Department of Agriculture, University of Brawijaya, Malang, East Java, Indonesia) 2(Department of Agriculture, University of Brawijaya, Malang, East Java, Indonesia) Abstract: Five Pandan species, namely Pandanus julianettii, Pandanus iwen, Pandanus brosimos, Pandanus sp.1 (owadak) and Pandanus sp.2 (woromo) from Jaya Wijaya Mountains, Papua have potential bioactive components. This study aimed to identify the bioactive components in Pandan’s fleshy receptacle and seed from five Pandan species. Samples were obtained from fleshy receptacle and seed. Sample were mashed and dried for further proximate, minerals, and vitamins contents analysis. The study revealed that nutritive value compositions of food fiber per 100 grams in fleshy receptacle and seed of Pandanus julianettii were 23% and 12%; those of Pandanus sp.1 (owadak) were 17.59% and 18.38%; those of Pandanus sp.2 (woromo) were 30% and 23%; those of Pandanus iwen were 18% and 30%; those of Pandanus brosimos were 47.75 % and 17.40%. The nutritive value compositions of starch per 100 grams in pandan’s fleshy receptacle and seed of Pandanus julianettii were 23 ppm and 0.24 ppm; those of Pandanus sp.1 (owadak) were 26 ppm and 0.96 ppm; those of Pandanus sp.2 (woromo) were 36 ppm and 18 ppm; those of Pandanus iwen were 21 ppm and 0.21 ppm; those of Pandanus brosimos were 35.88 ppm and 9.67 ppm. -

History of Agriculture in Papua New Guinea
History of agriculture in Papua New Guinea R. Michael Bourke sweet potato about 300 years ago; permanent settle- Introduction ment by Europeans and other outsiders, with many introductions of plants and animals after 1870; and the period of rapid social and economic change that The history of agriculture in PNG is about 10 000 commenced about 70 years ago in 1940. years old. This history is reviewed here in the context of 50 000 years of human occupation of the Australia – New Guinea region. 1 More is known about what The peopling of New Guinea has happened nearer to the present, especially since 1870, than about the distant past. Much of the early history (prehistory) of PNG was unknown until When the first humans came to New Guinea about about 50 years ago, but since 1959 there has been 50 000 years ago the climate was very different a lot of research on the prehistory of PNG, with a from now. Worldwide, temperatures were lower, major focus on agriculture. However, this is a rapidly the polar ice caps were larger, glaciers were more evolving field of study and our understanding of common, and sea levels were lower. As a result, the the history of agriculture in PNG is still incomplete. South-East Asia mainland extended as far east as The information that is summarised here will be Bali and Borneo to form a landmass that is known expanded and modified by future research. as Sunda. The Asian mainland (Sunda) and New Historical evidence is reviewed in a number of Guinea were always separated by ocean but, at that periods: the arrival of humans in New Guinea some time, New Guinea was not an island, but formed the 50 000 years ago; the beginnings of agriculture about northern part of a large continent that also included 10 000 years ago; the appearance of Austronesian- Australia and Tasmania, known as Sahul (Figure 1). -

Perennial Edible Fruits of the Tropics: an and Taxonomists Throughout the World Who Have Left Inventory
United States Department of Agriculture Perennial Edible Fruits Agricultural Research Service of the Tropics Agriculture Handbook No. 642 An Inventory t Abstract Acknowledgments Martin, Franklin W., Carl W. Cannpbell, Ruth M. Puberté. We owe first thanks to the botanists, horticulturists 1987 Perennial Edible Fruits of the Tropics: An and taxonomists throughout the world who have left Inventory. U.S. Department of Agriculture, written records of the fruits they encountered. Agriculture Handbook No. 642, 252 p., illus. Second, we thank Richard A. Hamilton, who read and The edible fruits of the Tropics are nnany in number, criticized the major part of the manuscript. His help varied in form, and irregular in distribution. They can be was invaluable. categorized as major or minor. Only about 300 Tropical fruits can be considered great. These are outstanding We also thank the many individuals who read, criti- in one or more of the following: Size, beauty, flavor, and cized, or contributed to various parts of the book. In nutritional value. In contrast are the more than 3,000 alphabetical order, they are Susan Abraham (Indian fruits that can be considered minor, limited severely by fruits), Herbert Barrett (citrus fruits), Jose Calzada one or more defects, such as very small size, poor taste Benza (fruits of Peru), Clarkson (South African fruits), or appeal, limited adaptability, or limited distribution. William 0. Cooper (citrus fruits), Derek Cormack The major fruits are not all well known. Some excellent (arrangements for review in Africa), Milton de Albu- fruits which rival the commercialized greatest are still querque (Brazilian fruits), Enriquito D. -

Des Vergers De Pandanus Spp. Comme Poste Avancé De La Culture Notule D’Ethnobotanique Baruya
Journal de la Société des Océanistes 114-115 | Année 2002 En hommage à Jacques Barrau Des vergers de Pandanus spp. comme poste avancé de la culture Notule d’ethnobotanique baruya Pierre Lemonnier Édition électronique URL : http://journals.openedition.org/jso/1480 DOI : 10.4000/jso.1480 ISSN : 1760-7256 Éditeur Société des océanistes Édition imprimée Date de publication : 1 décembre 2002 Pagination : 159-164 ISSN : 0300-953x Référence électronique Pierre Lemonnier, « Des vergers de Pandanus spp. comme poste avancé de la culture », Journal de la Société des Océanistes [En ligne], 114-115 | Année 2002, mis en ligne le 26 mai 2008, consulté le 22 mars 2021. URL : http://journals.openedition.org/jso/1480 ; DOI : https://doi.org/10.4000/jso.1480 Journal de la société des océanistes est mis à disposition selon les termes de la Licence Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de Modification 4.0 International. Des vergers de Pandanus spp. comme poste avancé de la culture. Notule d’ethnobotanique baruya par Pierre LEMONNIER* RÉSUMÉ ABSTRACT Chaque année, vers la mi-octobre, un quart de la Each year, towards mid-October, a quarter of the population baruya s’enfonce dans la forêt avec cochons, Baruya population heads up into the forest with their chiens et enfants, bien au-dessus des jardins les plus pigs, dogs and children, well above the highest cultivated élevés, pour collecter, surveiller et conserver les fruits gardens, to collect, watch over and preserve the fruits of d’arbres « semi-domestiqués » de la forêt, les Pandanus « semi-domesticated » forest trees, the Pandanus jiulia- jiulianettii et P. -

Papua New Guinea Papua New Guinea
COUNTRY REPORT ON THE STATE OF PLANT GENETIC RESOURCES FOR FOOD AND AGRICULTURE PAPUA NEW GUINEA PAPUA NEW GUINEA The State of Plant Genetic Resources for Food and Agriculture Country Report of Papua New Guinea September 2009 2 Note by FAO This Country Report has been prepared by the national authorities in the context of the preparatory process for the Second Report on the State of World’s Plant Genetic Resources for Food and Agriculture. The Report is being made available by the Food and Agriculture Organization of the United Nations (FAO) as requested by the Commission on Genetic Resources for Food and Agriculture. However, the report is solely the responsibility of the national authorities. The information in this report has not been verified by FAO, and the opinions expressed do not necessarily represent the views or policy of FAO. The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of FAO concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views of FAO. CONTENTS SECTION 1 -

Supplementary Materials Table S1. Plant Families That Include
Supplementary Materials Table S1. Plant families that include wild food plants (WFPs) and semi-cultivated species that are known to contribute to food and nutrition security in project countries and in the study cases mentioned in this review. Common Part Plant Family Species Name Range Name Eaten/Commercialized Amaranthus S. America, Amaranthaceae Amaranth Seeds, leaves tortuosus worldwide Chenopodium spp. Goosefoots Seeds Worldwide Ambacam, Anacardiaceae Mangifera foetida Fruit Southeast Asia Bacang * Foeniculum sp., cf Apiaceae Fennel Bulb, leaves Mediterranean F. vulgare South America— Apocynaceae Hancornia speciosa Mangaba * Fruits Brazil, Peru, Bolivia, Paraguay S. America—Bolivia, Astrocaryum Fruits, seeds, leaves, Brazil, Trinidad and Arecaceae Tucumã * aculeatum palm heart Tobago, Venezuela, Guyana, Suriname Wooly Jelly S. America—Southern Butia eriospatha Fruit and seed Palm Brazil Euterpe edulis Jussara * Fruit and palm heart Brazil Euterpe oleracea Açaí palm Fruit and seeds S. America—Brazil Eremurus Asphodelaceae Foxtail lily Leaves Mediterranean spectabilis Scolymus Asteraceae Golden thistle Root, young leaves Mediterranean hispanicus Porophyllum spp. Leaves Mesoamerica Diplazium Athyriaceae Vegetable fern Young leaves Asia, Oceania esculentum Nasturtium Brassicaceae Watercress Leaves Europe, Asia officinale Capparis spinosa Capparaceae Capers Fruits and flowers Mediterranean and C. decidua S. America—Brazil, Vasconcellea Bolivia, Peru, Caricaceae microcarpa (Carica Col de monte * Fruit, leaves Ecuador, Colombia, microcarpa) Venezuela; C. America—Panama Caryocar Caryocaraceae Pequi * Fruit, seeds Brazil brasiliense Worldwide in shallow Caulerpaceae Caulerpa racemosa Sea grapes Leaves temperate and tropical seas Cleomaceae Cleome gynandra Spider plant Leaves Africa Water spinach, Young shoots and Convolvulaceae Ipomoea aquatica water morning Southeast Asia leaves glory Garabato yuyo Dennstaedtiaceae Hypolepis hostilis Young shoots Amazonia * Tacca S.E. -

Agricultural Systems of Papua New Guinea
AUSTRALIAN AGENCY for INTERNATIONAL DEVELOPMENT AGRICULTURAL SYSTEMS OF PAPUA NEW GUINEA Working Paper No. 5 GULF PROVINCE TEXT SUMMARIES, MAPS, CODE LISTS AND VILLAGE IDENTIFICATION RL. Hide, R.M. Bourke, B.J. Allen, N. Fereday, D. Fritsch, R. Grau, E. Lowes and M. Woruba REVISED and REPRINTED 2002 THE AUSTRALIAN NA110NAL UNIVERSITY PAPUA NEW GUINEA DEPARTMENT OF AGRICULTURE AND LIVESTOCK UNIVERSITY OF.PAPUA NEW GUINEA AGRICULTURAL SYSTEMS OF PAPUA NEW GUINEA Working Paper No. 5 GULF PROVINCE TEXT SUMMARIES, MAPS, CODE LISTS AND VILLAGE IDENTIFICATION R.L. Hide, R.M. Bourke, B.J. Allen, N. Fereday, D. Fritsch, R. Grau, E. Lowes and M. Woruba Department of Human Geography, The Australian National University, ACT 0200, Australia REVISED and REPRINTED 2002 Correct Citation: Hide, R.L., Bourke, R.M., Allen, B.J., Fereday, N., Fritsch, D., Grau, R., Lowes, E. and Woruba, M. (2002). Gulf Province: Text Summaries, Maps, Code Lists and Village Identification. Agricultural Systems of Papua New Guinea Working Paper No. 5. Land Management Group, Department of Human Geography, Research School of Pacific and Asian Studies, The Australian National University, Canberra. Revised edition. National Library of Australia Cataloguing-in-Publication Entry: Gulf Province: text summaries, maps, code lists and village identification. Rev. ed. ISBN 0 9579381 5 2 1. Agricultural systems – Papua New Guinea – Gulf Province. 2. Agricultural geography – Papua New Guinea – Gulf Province. 3. Agricultural mapping – Papua New Guinea – Gulf Province. I. Hide, Robin Lamond. II. Australian National University. Land Management Group. (Series: Agricultural systems of Papua New Guinea working paper; no. 5). 630.99547 Cover Photograph: The late Gore Gabriel clearing undergrowth from a pandanus nut grove in the Sinasina area, Simbu Province (R.L. -

Food and Agriculture in Papua New Guinea
Food and Agriculture in Papua New Guinea edited by R. Michael Bourke and Tracy Harwood Published by ANU E Press The Australian National University Canberra ACT 0200, Australia Email: [email protected] This title is also available online at: http://epress.anu.edu.au/food_agriculture_citation.html This edition © 2009 ANU E Press All rights reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior permission of the publisher. National Library of Australia Cataloguing-in-Publication entry: Food and agriculture in Papua New Guinea / editors, R. Michael Bourke, Tracy Harwood. Bibliography. Includes index. ISBN 9781921536601 (pbk) ISBN 9781921536618 (pdf) Agriculture--Papua New Guinea Agriculture--Economic aspects--Papua New Guinea. Agricultural productivity--Papua New Guinea. 630.9953 Suggested citation: Bourke, R.M. and Harwood, T. (eds) (2009). Food and Agriculture in Papua New Guinea. ANU E Press, The Australian National University, Canberra. http://rspas.anu.edu.au/lmg/ Design: Sam Highley, Clarus Design Typeface: Minion Pro Index: Tracy Harwood Cover: Yasisime from Asirangka village in the Aiyura basin, Eastern Highlands Province, weeds a peanut plot. Peanut is grown for both home consumption and for sale locally. It is grown in rotation with sweet potato to maintain soil fertility. The plot on the left contains sweet potato, the staple food in the highlands, and other food crops. Casuarina trees in the background shade Arabica coffee, the most important cash crop in the highlands. Photo by Mike Bourke. Contents Foreword . vii Charles W. -
Environment and Food Production in Papua New Guinea
4 Environment and Food Production in Papua New Guinea R. Michael Bourke Introduction The physical environment plays a critical role in determining both where crops can be grown and how productive they are. The environment in which plants grow can be modified to some degree by people, for example, using drainage to reduce excessive soil moisture. Here the main physical environmental factors that influence crop growth in Papua New Guinea (PNG) are noted and their influence on crop distribution and productivity is reviewed. This research builds on the pioneering work of Harold Brookfield (1964) on the ecology of human settlement in New Guinea over half a century ago. The focus here is on those crops that were used in PNG before the recent adoption of numerous introduced species. Most of these new crops appeared after 1870 following sustained European contact, but sweet potato came into PNG via Southeast Asia about 300 years ago (Ballard 2005: 9; Bayliss-Smith et al. 2005: 109). Tobacco, another American plant, was present in west New Guinea about 400 years ago (Ballard 2005: 8). I pay more attention to the impact of temperature as small differences can have a significant influence on both the distribution and productivity of crops, particularly near their environmental margins. Greater emphasis is also given to the PNG highlands than to the lowlands or the intermediate zone because the focus of this volume is on the Kuk archaeological site and the Wahgi Valley in the highlands. The environmental factors considered here, besides temperature, are soil moisture (including flooding), cloud cover, slope, day length and soil fertility. -
List of Culinary Nuts
List of culinary nuts • Gymnosperm seeds: naked seeds, with no enclosure (e.g. pine nuts); • Angiosperm seeds: unenclosed seeds within a larger fruit (e.g. peanuts). Nuts have a rich history as food. For many indigenous peoples of the Americas, a wide variety of nuts, includ- ing acorns, American beech, and others, served as a ma- jor source of starch and fat over thousands of years.[5] Similarly, a wide variety of nuts have served as food for Indigenous Australians for many centuries.[6] Other culinary nuts, though known from ancient times, have seen dramatic increases in use in modern times. The most striking such example is the peanut. Its usage was popularized by the work of George Washington Carver, A small bowl of mixed nuts who discovered and popularized many applications of the peanut after employing peanut plants for soil ameliora- tion in fields used to grow cotton.[7] 1 True nuts A typical assortment of mixed nuts Culinary nuts are dry, edible fruits or seeds that usually, but not always, have a high fat content. Nuts are used in a wide variety of edible roles, including in baking, as The common hazelnut, native to Europe and Western Asia snacks (either roasted or raw), and as flavoring. In addi- tion to botanical nuts, fruits and seeds that have a similar The following are both culinary and botanical nuts. appearance and culinary role are considered to be culi- nary nuts.[1] Culinary nuts are divided into fruits or seeds in one of four categories: • Acorn (Quercus, Lithocarpus and Cyclobalanopsis spp.), used from ancient times among indigenous peoples of the Americas as a staple food, in partic- • True, or botanical nuts: dry, hard-shelled, uncom- ular for making bread and porridge.[8] partmented fruit that do not split on maturity to re- lease seeds;[2][3] • Beech (Fagus spp.) • Drupes: fleshy fruit surrounding a stone, or pit, con- • American beech (Fagus grandifolia), used by taining a seed (e.g.