Male Mating Behaviour and Mating Systems of Bees: an Overview Robert John Paxton
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Barrett, M., S. Schneider, P. Sachdeva, A. Gomez, S. Buchmann
Journal of Comparative Physiology A https://doi.org/10.1007/s00359-021-01492-4 ORIGINAL PAPER Neuroanatomical diferentiation associated with alternative reproductive tactics in male arid land bees, Centris pallida and Amegilla dawsoni Meghan Barrett1 · Sophi Schneider2 · Purnima Sachdeva1 · Angelina Gomez1 · Stephen Buchmann3,4 · Sean O’Donnell1,5 Received: 1 February 2021 / Revised: 19 May 2021 / Accepted: 22 May 2021 © The Author(s), under exclusive licence to Springer-Verlag GmbH Germany, part of Springer Nature 2021 Abstract Alternative reproductive tactics (ARTs) occur when there is categorical variation in the reproductive strategies of a sex within a population. These diferent behavioral phenotypes can expose animals to distinct cognitive challenges, which may be addressed through neuroanatomical diferentiation. The dramatic phenotypic plasticity underlying ARTs provides a powerful opportunity to study how intraspecifc nervous system variation can support distinct cognitive abilities. We hypothesized that conspecifc animals pursuing ARTs would exhibit dissimilar brain architecture. Dimorphic males of the bee species Centris pallida and Amegilla dawsoni use alternative mate location strategies that rely primarily on either olfaction (large-morph) or vision (small-morph) to fnd females. This variation in behavior led us to predict increased volumes of the brain regions supporting their primarily chemosensory or visual mate location strategies. Large-morph males relying mainly on olfaction had relatively larger antennal lobes and relatively smaller optic lobes than small-morph males relying primarily on visual cues. In both species, as relative volumes of the optic lobe increased, the relative volume of the antennal lobe decreased. In addition, A. dawsoni large males had relatively larger mushroom body lips, which process olfactory inputs. -

Historia Natural De Macrotera Pipiyolin (Hymenoptera: Andrenidae) En La Estación De Biología Chamela, Jalisco, México
Revista Mexicana de Biodiversidad 81: 87- 96, 2010 http://dx.doi.org/10.22201/ib.20078706e.2010.001.196 Historia natural de Macrotera pipiyolin (Hymenoptera: Andrenidae) en la Estación de Biología Chamela, Jalisco, México Natural history of Macrotera pipiyolin (Hymenoptera: Andrenidae) in the Estación de Biología Chamela, Jalisco, Mexico Beatriz Rodríguez-Velez1* y Ricardo Ayala2 1Instituto de Biología, Departamento de Zoología, Universidad Nacional Autónoma de México. Apartado postal 70-153, 04510 México, D.F., México. 2Estación de Biología Chamela, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado postal 21, San Patricio, 48980 Jalisco, México. *Correspondencia: [email protected] Resumen. Se presenta información sobre la historia natural de Macrotera pipiyolin; el estudio se realizó en la Estación de Biología Chamela, Instituto de Biología, UNAM, entre junio y agosto de 1994. Las abejas emergieron poco después del inicio de la temporada de lluvias, su pico de actividad fue a principio de julio, presentando alrededor de 35 días activos. La actividad concuerda con la fl oración de Opuntia excelsa, planta de la que obtiene sus recursos. Las cópulas ocurren sobre las fl ores y los machos muestran 3 tipos de estrategias reproductivas: macho dominante, subordinado y patrullero. Las hembras son poliándricas y los machos polígamos. Las hembras construyen los nidos generalmente cerca de O. excelsa y tienen una distribución gregaria. Los nidos son simples, con un túnel principal y celdas, las que se cierran y desconectan del tunel una vez que se han aprovisionado y en ellas se ha depositado un huevo. Para la provisión de una celda se requiere de 2 a 18 cargas de polen. -

Food Load Manipulation Ability Shapes Flight Morphology in Females Of
Polidori et al. Frontiers in Zoology 2013, 10:36 http://www.frontiersinzoology.com/content/10/1/36 RESEARCH Open Access Food load manipulation ability shapes flight morphology in females of central-place foraging Hymenoptera Carlo Polidori1*, Angelica Crottini2, Lidia Della Venezia3,5, Jesús Selfa4, Nicola Saino5 and Diego Rubolini5 Abstract Background: Ecological constraints related to foraging are expected to affect the evolution of morphological traits relevant to food capture, manipulation and transport. Females of central-place foraging Hymenoptera vary in their food load manipulation ability. Bees and social wasps modulate the amount of food taken per foraging trip (in terms of e.g. number of pollen grains or parts of prey), while solitary wasps carry exclusively entire prey items. We hypothesized that the foraging constraints acting on females of the latter species, imposed by the upper limit to the load size they are able to transport in flight, should promote the evolution of a greater load-lifting capacity and manoeuvrability, specifically in terms of greater flight muscle to body mass ratio and lower wing loading. Results: Our comparative study of 28 species confirms that, accounting for shared ancestry, female flight muscle ratio was significantly higher and wing loading lower in species taking entire prey compared to those that are able to modulate load size. Body mass had no effect on flight muscle ratio, though it strongly and negatively co-varied with wing loading. Across species, flight muscle ratio and wing loading were negatively correlated, suggesting coevolution of these traits. Conclusions: Natural selection has led to the coevolution of resource load manipulation ability and morphological traits affecting flying ability with additional loads in females of central-place foraging Hymenoptera. -
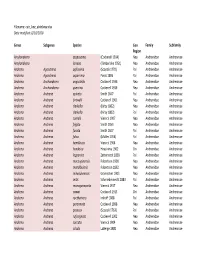
Anthophila List
Filename: cuic_bee_database. -

Molecular Ecology and Social Evolution of the Eastern Carpenter Bee
Molecular ecology and social evolution of the eastern carpenter bee, Xylocopa virginica Jessica L. Vickruck, B.Sc., M.Sc. Department of Biological Sciences Submitted in partial fulfillment of the requirements for the degree of PhD Faculty of Mathematics and Science, Brock University St. Catharines, Ontario © 2017 Abstract Bees are extremely valuable models in both ecology and evolutionary biology. Their link to agriculture and sensitivity to climate change make them an excellent group to examine how anthropogenic disturbance can affect how genes flow through populations. In addition, many bees demonstrate behavioural flexibility, making certain species excellent models with which to study the evolution of social groups. This thesis studies the molecular ecology and social evolution of one such bee, the eastern carpenter bee, Xylocopa virginica. As a generalist native pollinator that nests almost exclusively in milled lumber, anthropogenic disturbance and climate change have the power to drastically alter how genes flow through eastern carpenter bee populations. In addition, X. virginica is facultatively social and is an excellent organism to examine how species evolve from solitary to group living. Across their range of eastern North America, X. virginica appears to be structured into three main subpopulations: a northern group, a western group and a core group. Population genetic analyses suggest that the northern and potentially the western group represent recent range expansions. Climate data also suggest that summer and winter temperatures describe a significant amount of the genetic differentiation seen across their range. Taken together, this suggests that climate warming may have allowed eastern carpenter bees to expand their range northward. Despite nesting predominantly in disturbed areas, eastern carpenter bees have adapted to newly available habitat and appear to be thriving. -

Components of Nest Provisioning Behavior in Solitary Bees (Hymenoptera: Apoidea)*
Apidologie 39 (2008) 30–45 Available online at: c INRA/DIB-AGIB/ EDP Sciences, 2008 www.apidologie.org DOI: 10.1051/apido:2007055 Review article Components of nest provisioning behavior in solitary bees (Hymenoptera: Apoidea)* John L. Neff Central Texas Melittological Institute, 7307 Running Rope, Austin, Texas 78731, USA Received 13 June 2007 – Revised 28 September 2007 – Accepted 1 October 2007 Abstract – The components of nest provisioning behavior (resources per cell, transport capacity, trip dura- tion, trips per cell) are examined for a data set derived from the literature and various unpublished studies. While there is a trade-off between transport capacity and trips required per cell, the highest provisioning rates are achieved by bees carrying very large pollen loads at intermediate trip durations. Most solitary bees appear to be either egg or resource limited, so sustained provisioning rates over one cell per day are unusual. Provisioning rate / body size / transport capacity / solitary bees / trip duration 1. INTRODUCTION tradeoffs between provisioning one large cell or two small cells direct, indirect, or nonex- There is a vast literature on the foraging tac- istent? Are provisioning rates related to body tics of bees: what kinds of flowers to visit, how size? long to spend on a flower, how many flowers Provisioning rate is a general term and can in an inflorescence to visit, when to turn, and simply refer to the rate (mass per unit time) so forth. Much of this work is done within a at which foragers bring various food items framework of whether a forager should max- (pollen, nectar, oils, carrion, or whatever) to imize harvesting rate or foraging efficiency their nests. -

The Impact of Molecular Data on Our Understanding of Bee Phylogeny and Evolution
EN58CH04-Danforth ARI 5 December 2012 7:55 The Impact of Molecular Data on Our Understanding of Bee Phylogeny and Evolution Bryan N. Danforth,1∗ Sophie Cardinal,2 Christophe Praz,3 Eduardo A.B. Almeida,4 and Denis Michez5 1Department of Entomology, Cornell University, Ithaca, New York 14853; email: [email protected] 2Canadian National Collection of Insects, Agriculture Canada, Ottawa, Ontario K1A 0C6, Canada; email: [email protected] 3Institute of Biology, University of Neuchatel, Emile-Argand 11, 2009 Neuchatel, Switzerland; email: [email protected] 4Departamento de Biologia, FFCLRP-Universidade de Sao˜ Paulo, 14040-901 Ribeirao˜ Preto, Sao˜ Paulo, Brazil; email: [email protected] 5University of Mons, Laboratory of Zoology, 7000 Mons, Belgium; email: [email protected] Annu. Rev. Entomol. 2013. 58:57–78 Keywords First published online as a Review in Advance on Hymenoptera, Apoidea, bees, molecular systematics, sociality, parasitism, August 28, 2012 plant-insect interactions The Annual Review of Entomology is online at ento.annualreviews.org Abstract by 77.56.160.109 on 01/14/13. For personal use only. This article’s doi: Our understanding of bee phylogeny has improved over the past fifteen years 10.1146/annurev-ento-120811-153633 as a result of new data, primarily nucleotide sequence data, and new methods, Copyright c 2013 by Annual Reviews. primarily model-based methods of phylogeny reconstruction. Phylogenetic All rights reserved Annu. Rev. Entomol. 2013.58:57-78. Downloaded from www.annualreviews.org studies based on single or, more commonly, multilocus data sets have helped ∗ Corresponding author resolve the placement of bees within the superfamily Apoidea; the relation- ships among the seven families of bees; and the relationships among bee subfamilies, tribes, genera, and species. -

Insects and Related Arthropods Associated with of Agriculture
USDA United States Department Insects and Related Arthropods Associated with of Agriculture Forest Service Greenleaf Manzanita in Montane Chaparral Pacific Southwest Communities of Northeastern California Research Station General Technical Report Michael A. Valenti George T. Ferrell Alan A. Berryman PSW-GTR- 167 Publisher: Pacific Southwest Research Station Albany, California Forest Service Mailing address: U.S. Department of Agriculture PO Box 245, Berkeley CA 9470 1 -0245 Abstract Valenti, Michael A.; Ferrell, George T.; Berryman, Alan A. 1997. Insects and related arthropods associated with greenleaf manzanita in montane chaparral communities of northeastern California. Gen. Tech. Rep. PSW-GTR-167. Albany, CA: Pacific Southwest Research Station, Forest Service, U.S. Dept. Agriculture; 26 p. September 1997 Specimens representing 19 orders and 169 arthropod families (mostly insects) were collected from greenleaf manzanita brushfields in northeastern California and identified to species whenever possible. More than500 taxa below the family level wereinventoried, and each listing includes relative frequency of encounter, life stages collected, and dominant role in the greenleaf manzanita community. Specific host relationships are included for some predators and parasitoids. Herbivores, predators, and parasitoids comprised the majority (80 percent) of identified insects and related taxa. Retrieval Terms: Arctostaphylos patula, arthropods, California, insects, manzanita The Authors Michael A. Valenti is Forest Health Specialist, Delaware Department of Agriculture, 2320 S. DuPont Hwy, Dover, DE 19901-5515. George T. Ferrell is a retired Research Entomologist, Pacific Southwest Research Station, 2400 Washington Ave., Redding, CA 96001. Alan A. Berryman is Professor of Entomology, Washington State University, Pullman, WA 99164-6382. All photographs were taken by Michael A. Valenti, except for Figure 2, which was taken by Amy H. -

The Very Handy Bee Manual
The Very Handy Manual: How to Catch and Identify Bees and Manage a Collection A Collective and Ongoing Effort by Those Who Love to Study Bees in North America Last Revised: October, 2010 This manual is a compilation of the wisdom and experience of many individuals, some of whom are directly acknowledged here and others not. We thank all of you. The bulk of the text was compiled by Sam Droege at the USGS Native Bee Inventory and Monitoring Lab over several years from 2004-2008. We regularly update the manual with new information, so, if you have a new technique, some additional ideas for sections, corrections or additions, we would like to hear from you. Please email those to Sam Droege ([email protected]). You can also email Sam if you are interested in joining the group’s discussion group on bee monitoring and identification. Many thanks to Dave and Janice Green, Tracy Zarrillo, and Liz Sellers for their many hours of editing this manual. "They've got this steamroller going, and they won't stop until there's nobody fishing. What are they going to do then, save some bees?" - Mike Russo (Massachusetts fisherman who has fished cod for 18 years, on environmentalists)-Provided by Matthew Shepherd Contents Where to Find Bees ...................................................................................................................................... 2 Nets ............................................................................................................................................................. 2 Netting Technique ...................................................................................................................................... -

The Morphology and Behavior of Dimorphic Males in Perdita Portalis
Behav Ecol Sociobiol (1991) 29:235-247 Behavioral Ecology and Sociobiology © Springer-Verlag 1991 The morphology and behavior of dimorphic males in Perdita portMis (Hymenoptera: Andrenidae) Bryan N. Danforth* Snow Entomological Museum, Department of Entomology, University of Kansas, Lawrence, KS 66045, USA Received September 3, 1990 / Accepted June 4, 1991 Summary. In Perdita portalis, a ground nesting, commu- found on flowers visited by foraging females (SH nal bee, males are clearly dimorphic. The two male morph). First discovered by Rozen (1970), the other, morphs are easily distinguished based on head size and derived, morph is strikingly modified, as judged by com- shape into (1) a flight-capable, small-headed (SH) morph parison with related bees, possessing an expanded head that resembles the males of other closely related species capsule, enlarged facial foveae, reduced compound eyes, and (2) a flightless, large-headed (LH) morph that pos- elongate pointed mandibles, a three-pronged clypeus, sesses numerous derived traits, such as reduced com- and reduced thoracic musculature, which renders it pound eyes, enlarged facial foveae and fully atrophied flightless and restricted to a life within the natal nest indirect flight muscles. The SH morph occurs exclusively (LH morph). Unlike most of the close relatives of Perdita on flowers while the LH morph is found only in nests portalis, which show continuous variation in head size, with females. While on flowers, SH males are aggressive, albeit with strong positive allometry, the males of P. fighting with conspecific males and heterospecific male portalis are truly dimorphic. and female bees, and they mate frequently with foraging Flightless, macrocephalic males are known from a females. -

Melissa 6, January 1993
The Melittologist's Newsletter Ronald J. McGinley. Bryon N. Danforth. Maureen J. Mello Deportment of Entomology • Smithsonian Institution. NHB-105 • Washington. DC 20560 NUMBER-6 January, 1993 CONTENTS COLLECTING NEWS COLLECTING NEWS .:....:Repo=.:..:..rt=on~Th.:..:.=ird=-=-PC=A..::.;M:.:...E=xp=ed=it=io:..o..n-------=-1 Report on Third PCAM Expedition Update on NSF Mexican Bee Inventory 4 Robert W. Brooks ..;::.LC~;.,;;;....;;...:...:...;....;...;;;..o,__;_;.c..="-'-'-;.;..;....;;;~.....:.;..;.""""""_,;;...;....________,;. Snow Entomological Museum .:...P.:...roposo.a:...::..;:;..,:=..ed;::....;...P_:;C"'-AM~,;::S,;::u.;...;rvc...;;e.L.y-'-A"'"-rea.:o..=s'------------'-4 University of Kansas Lawrence, KS 66045 Collecting on Guana Island, British Virgin Islands & Puerto Rico 5 The third NSF funded PCAM (Programa Cooperativo so- RESEARCH NEWS bre Ia Apifauna Mexicana) expedition took place from March 23 to April3, 1992. The major goals of this trip were ...:..T.o..:he;::....;...P.:::a::.::ra::.::s;:;;it:..::ic;....;;B::...:e:..:e:....::L=.:e:.:.ia;:;L'{XJd:..::..::..::u;,;;:.s....:::s.:..:.in.:..o~gc::u.:.::/a:o.:n;,;;:.s_____~7 to do springtime collecting in the Chihuahuan Desert and Decline in Bombus terrestris Populations in Turkey 7 Coahuilan Inland Chaparral habitats of northern Mexico. We =:...:::=.:.::....:::..:...==.:.:..:::::..:::.::...:.=.:..:..:=~:....=..~==~..:.:.:.....:..=.:=L-.--=- also did some collecting in coniferous forest (pinyon-juni- NASA Sponsored Solitary Bee Research 8 per), mixed oak-pine forest, and riparian habitats in the Si- ;:...;N:..::.o=tes;.,;;;....;o;,;,n:....:Nc..:.e.::..;st;:.;,i;,:,;n_g....:::b""-y....:.M=-'-e.;;,agil,;;a.;_:;c=h:..:..:ili=d-=B;....;;e....:::e..::.s______.....:::.8 erra Madre Oriental. Hymenoptera Database System Update 9 Participants in this expedition were Ricardo Ayala (Insti- '-'M:.Liss.:..:..:..;;;in..:....:g..;:JB<:..;ee:..::.::.::;.:..,;Pa:::..rt=s=?=::...;:_"'-L..;=c.:.:...c::.....::....::=:..,_----__;:_9 tuto de Biologia, Chamela, Jalisco); John L. -
Kennedy Range National Park and Proposed Additions 2008 Management Plan No 59 CONTENTS PART A
Kennedy Range National Park and Proposed Additions 2008 Management Plan No 59 CONTENTS PART A. INTRODUCTION PART A. INTRODUCTION. .1 1. Brief Overview. .1 1. BRIEF Overview 2. Key Values. 1 Kennedy Range National Park is located approximately 150 km PART B. MANAGEMENT DIRECTIONS AND PURPOSE. 3 east of Carnarvon and approximately 15 km north of Gascoyne 3. Vision. 3 Junction. The park and its proposed additions encompass 4. Legislative Framework . 3 319 037 hectares. 5. Management Arrangements with Indigenous People . 4 6. Existing and Proposed Tenure . 4 The range is a remarkable landscape feature which rises about 7. Performance Assessment. .5 100 m above the surrounding plain and comprises an isolated 8. Naming of Sites and Features. .5 remnant of an older land surface. Apart from its outstanding geology and scenic beauty, the park is valued for a variety of PART C. MANAGING THE NATURAL ENVIRONMENT. 6 natural values. 9. Biogeography. 6 10. Wilderness. 6 The park is located within the Western Australian Planning 11. Climate and Climate Change . .8 Commission’s (WAPC) Gascoyne Planning Region of Western 12. Geology, Geomorphology and Land Systems. .9 Australia and within the Shires of Carnarvon and Upper 13. Hydrology and Catchment Protection. 12 Gascoyne. 14. Native Plants and Plant Communities. 13 15. Native Animals and Habitats. .16 The proposed additions of 177 377 ha were purchased with 16. Threatened Ecological Communities. 20 the intention to add the area to the public conservation estate 17. Environmental Weeds. 20 (nominally as national park). The purchases comprise the Mooka 18. Introduced and Other Problem Animals .