This Electronic Thesis Or Dissertation Has Been Downloaded from Explore Bristol Research
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Methods and Work Profile
REVIEW OF THE KNOWN AND POTENTIAL BIODIVERSITY IMPACTS OF PHYTOPHTHORA AND THE LIKELY IMPACT ON ECOSYSTEM SERVICES JANUARY 2011 Simon Conyers Kate Somerwill Carmel Ramwell John Hughes Ruth Laybourn Naomi Jones Food and Environment Research Agency Sand Hutton, York, YO41 1LZ 2 CONTENTS Executive Summary .......................................................................................................................... 8 1. Introduction ............................................................................................................ 13 1.1 Background ........................................................................................................................ 13 1.2 Objectives .......................................................................................................................... 15 2. Review of the potential impacts on species of higher trophic groups .................... 16 2.1 Introduction ........................................................................................................................ 16 2.2 Methods ............................................................................................................................. 16 2.3 Results ............................................................................................................................... 17 2.4 Discussion .......................................................................................................................... 44 3. Review of the potential impacts on ecosystem services ....................................... -

The Digger Wasps of Saudi Arabia: New Records and Distribution, with a Checklist of Species (Hym.: Ampulicidae, Crabronidae and Sphecidae)
NORTH-WESTERN JOURNAL OF ZOOLOGY 9 (2): 345-364 ©NwjZ, Oradea, Romania, 2013 Article No.: 131206 http://biozoojournals.3x.ro/nwjz/index.html The digger wasps of Saudi Arabia: New records and distribution, with a checklist of species (Hym.: Ampulicidae, Crabronidae and Sphecidae) Neveen S. GADALLAH1,*, Hathal M. AL DHAFER2, Yousif N. ALDRYHIM2, Hassan H. FADL2 and Ali A. ELGHARBAWY2 1. Entomology Department, Faculty of Science, Cairo University, Giza, Egypt. 2. Plant Protection Department, College of Food and Agriculture Science, King Saud University, King Saud Museum of Arthropod (KSMA), Riyadh, Saudi Arabia. *Corresponing author, N.S. Gadalah, E-mail: [email protected] Received: 24. September 2012 / Accepted: 13. January 2013 / Available online: 02. June 2013 / Printed: December 2013 Abstract. The “sphecid’ fauna of Saudi Arabia (Hymenoptera: Apoidea) is listed. A total of 207 species in 42 genera are recorded including previous and new species records. Most Saudi Arabian species recorded up to now are more or less common and widespread mainly in the Afrotropical and Palaearctic zoogeographical zones, the exception being Bembix buettikeri Guichard, Bembix hofufensis Guichard, Bembix saudi Guichard, Cerceris constricta Guichard, Oxybelus lanceolatus Gerstaecker, Palarus arabicus Pulawski in Pulawski & Prentice, Tachytes arabicus Guichard and Tachytes fidelis Pulawski, which are presumed endemic to Saudi Arabia (3.9% of the total number of species). General distribution and ecozones, and Saudi Arabian localities are given for each species. In this study two genera (Diodontus Curtis and Dryudella Spinola) and 11 species are newly recorded from Saudi Arabia. Key words: Ampulicidae, Crabronidae, Sphecidae, faunistic list, new records, Saudi Arabia. Introduction tata boops (Schrank), Bembecinus meridionalis A.Costa, Diodontus sp. -

Coversheet for Thesis in Sussex Research Online
A University of Sussex DPhil thesis Available online via Sussex Research Online: http://sro.sussex.ac.uk/ This thesis is protected by copyright which belongs to the author. This thesis cannot be reproduced or quoted extensively from without first obtaining permission in writing from the Author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the Author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given Please visit Sussex Research Online for more information and further details Information gathering and conflict resolution in Polistes wasps Jonathan Philip Green Submitted for the degree of Doctor of Philosophy University of Sussex September 2011 ii Declaration The design and data collection for the study presented in Chapter 4 were undertaken in collaboration with Dr. Elli Leadbeater at the Institute of Zoology and Professor Jeremy Field at the University of Sussex. However, the particular analyses undertaken in that chapter, as well as the interpretations drawn from the data, are my own. I certify that, with the above qualification, the work carried out in this thesis is entirely my own, and that any help provided by other individuals with data collection and analysis is fully acknowledged. In addition, I certify that this thesis has not been, and will not be, submitted in whole or in part to another university for the award of any other degree. Signature: Jonathan Philip Green iii UNIVERISTY OF SUSSEX JONATHAN PHILIP GREEN, DOCTOR OF PHILOSOPHY INFORMATION GATHERING AND CONFLICT RESOLUTION IN POLISTES WASPS SUMMARY Signals are used to communicate resource-holding potential (RHP) to rivals during contests across a wide range of taxa. -

Lepidoptera on the Introduced Robinia Pseudoacacia in Slovakia, Central Europe
Check List 8(4): 709–711, 2012 © 2012 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution Lepidoptera on the introduced Robinia pseudoacacia in PECIES S OF ISTS L Slovakia, Central Europe Miroslav Kulfan E-mail: [email protected] Comenius University, Faculty of Natural Sciences, Department of Ecology, Mlynská dolina B-1, SK-84215 Bratislava, Slovakia. Abstract: Robinia pseudoacacia A current checklist of Lepidoptera that utilize as a hostplant in Slovakia (Central Europe) faunalis provided. community. The inventory Two monophagous is based on species, a bibliographic the leaf reviewminers andMacrosaccus new unreported robiniella data and from Parectopa southwest robiniella Slovakia., and Thethe polyphagouslist includes 35pest Lepidoptera Hyphantria species cunea belonging to 10 families. Most species are polyphagous and belong to Euro-Siberian have subsequently been introduced to Slovakia. Introduction E. The area is a polygon enclosed by the towns of Bratislava, Robinia pseudoacacia a widespread species in its native habitat in southeastern North America. It was L.introduced (black locust, to orEurope false acacia),in 1601 is Komárno, Veľký Krtíš and Myjava. Ten plots were located in the southern part of the study area. Most were located in theThe remnant trophic ofgroups the original of the floodplain Lepidoptera forests larvae that found were (Chapman 1935). The first mention of planting the species distributed along the Danube and Morava rivers. (Keresztesiin Slovakia dates 1965). from Today, 1750, itwhen is widespread black locust wasthroughout planted (1986). The zoogeographical distribution of the species western,around the central, fortress eastern in Komárno and southern in southern Europe, Slovakia where followswere defined the arrangement following the give system by Reiprichof Brown (2001). -

Systematics of Polistes (Hymenoptera: Vespidae), with a Phylogenetic Consideration of Hamilton’S Haplodiploidy Hypothesis
Ann. Zool. Fennici 43: 390–406 ISSN 0003-455X Helsinki 29 December 2006 © Finnish Zoological and Botanical Publishing Board 2006 Systematics of Polistes (Hymenoptera: Vespidae), with a phylogenetic consideration of Hamilton’s haplodiploidy hypothesis Kurt M. Pickett*, James M. Carpenter & Ward C. Wheeler Division of Invertebrate Zoology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10023, USA * Current address: Department of Biology, University of Vermont, Room 120A Marsh Life Science Building, 109 Carrigan Drive, Burlington, VT 05405, USA Received 30 Nov. 2005, revised version received 21 Nov. 2006, accepted 4 May 2006 Pickett, K. M., Carpenter, J. M. & Wheeler, W. C. 2006: Systematics of Polistes (Hymenoptera: Vespidae), with a phylogenetic consideration of Hamilton’s haplodiploidy hypothesis. — Ann. Zool. Fennici 43: 390–406. A review of previously published cladistic analyses of Polistes is presented. The two most recent analyses of Polistes are shown to be largely consistent phylogenetically. Although the taxonomy implied by each differs, this difference is shown to be mostly due to taxon sampling. After the review, a phylogenetic analysis of Polistes — the most data-rich yet undertaken — is presented. The analysis includes new data and the data from previously published analyses. The differing conclusions of the previous studies are discussed in light of the new analysis. After discussing the status of subge- neric taxonomy in Polistes, the new phylogeny is used to test an important hypothesis regarding the origin of social behavior: the haplodiploidy hypothesis of Hamilton. Prior phylogenetic analyses so while these studies achieved their goal, with within Polistes resolutions leading to rejection of Emery’s Rule, they had little to say about broader phylogenetic Cladistic analysis of species-level relationships patterns within the genus. -

Hymenoptera: Vespoidea) for the Colombian Orinoco Region Biota Colombiana, Vol
Biota Colombiana ISSN: 0124-5376 ISSN: 2539-200X [email protected] Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt" Colombia Halmenschlager, Matheus; Agudelo Martínez, Juan C; Pérez-Buitrago, Néstor F. New records of Vespidae (Hymenoptera: Vespoidea) for the Colombian Orinoco Region Biota Colombiana, vol. 20, no. 1, 2019, January-June, pp. 21-33 Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt" Colombia Available in: https://www.redalyc.org/articulo.oa?id=49159822002 How to cite Complete issue Scientific Information System Redalyc More information about this article Network of Scientific Journals from Latin America and the Caribbean, Spain and Journal's webpage in redalyc.org Portugal Project academic non-profit, developed under the open access initiative Halmenschlager et al. New records of wasps in the Colombian Orinoco New records of Vespidae (Hymenoptera: Vespoidea) for the Colombian Orinoco Region Nuevos registros de Vespidae (Hymenoptera: Vespoidea) para la región de la Orinoquía colombiana Matheus Y. Halmenschlager, Juan C. Agudelo Martínez and Néstor F. Pérez-Buitrago Abstract We analyzed 72 specimens from the Arauca (71) and Casanare (1) departments in the Orinoco region of Colombia. 7KHVSHFLPHQVEHORQJWRJHQHUDDQGVSHFLHVRIYHVSLGZDVSV)RXUVSHFLHVDUHUHSRUWHGIRUWKHÀUVWWLPH for the region and 14 are new records for the Arauca department. There is a likely new record of Stenodynerus cf. australis for the Neotropical region. Keywords. Arauca. Eumeninae. Neotropic. Species list. Vespid wasps. Resumen Analizamos 72 especímenes colectados de los departamentos de Arauca (71) y Casanare (1) en la región de la Orinoquía. Estos pertenecen a 10 géneros y 18 especies de avispas. Cuatro especies son nuevos registros para la región y 14 son nuevas para el departamento de Arauca. -

Island Biology Island Biology
IIssllaanndd bbiioollooggyy Allan Sørensen Allan Timmermann, Ana Maria Martín González Camilla Hansen Camille Kruch Dorte Jensen Eva Grøndahl, Franziska Petra Popko, Grete Fogtmann Jensen, Gudny Asgeirsdottir, Hubertus Heinicke, Jan Nikkelborg, Janne Thirstrup, Karin T. Clausen, Karina Mikkelsen, Katrine Meisner, Kent Olsen, Kristina Boros, Linn Kathrin Øverland, Lucía de la Guardia, Marie S. Hoelgaard, Melissa Wetter Mikkel Sørensen, Morten Ravn Knudsen, Pedro Finamore, Petr Klimes, Rasmus Højer Jensen, Tenna Boye Tine Biedenweg AARHUS UNIVERSITY 2005/ESSAYS IN EVOLUTIONARY ECOLOGY Teachers: Bodil K. Ehlers, Tanja Ingversen, Dave Parker, MIchael Warrer Larsen, Yoko L. Dupont & Jens M. Olesen 1 C o n t e n t s Atlantic Ocean Islands Faroe Islands Kent Olsen 4 Shetland Islands Janne Thirstrup 10 Svalbard Linn Kathrin Øverland 14 Greenland Eva Grøndahl 18 Azores Tenna Boye 22 St. Helena Pedro Finamore 25 Falkland Islands Kristina Boros 29 Cape Verde Islands Allan Sørensen 32 Tristan da Cunha Rasmus Højer Jensen 36 Mediterranean Islands Corsica Camille Kruch 39 Cyprus Tine Biedenweg 42 Indian Ocean Islands Socotra Mikkel Sørensen 47 Zanzibar Karina Mikkelsen 50 Maldives Allan Timmermann 54 Krakatau Camilla Hansen 57 Bali and Lombok Grete Fogtmann Jensen 61 Pacific Islands New Guinea Lucía de la Guardia 66 2 Solomon Islands Karin T. Clausen 70 New Caledonia Franziska Petra Popko 74 Samoa Morten Ravn Knudsen 77 Tasmania Jan Nikkelborg 81 Fiji Melissa Wetter 84 New Zealand Marie S. Hoelgaard 87 Pitcairn Katrine Meisner 91 Juan Fernandéz Islands Gudny Asgeirsdottir 95 Hawaiian Islands Petr Klimes 97 Galápagos Islands Dorthe Jensen 102 Caribbean Islands Cuba Hubertus Heinicke 107 Dominica Ana Maria Martin Gonzalez 110 Essay localities 3 The Faroe Islands Kent Olsen Introduction The Faroe Islands is a treeless archipelago situated in the heart of the warm North Atlantic Current on the Wyville Thompson Ridge between 61°20’ and 62°24’ N and between 6°15’ and 7°41’ W. -

Historia Natural De Macrotera Pipiyolin (Hymenoptera: Andrenidae) En La Estación De Biología Chamela, Jalisco, México
Revista Mexicana de Biodiversidad 81: 87- 96, 2010 http://dx.doi.org/10.22201/ib.20078706e.2010.001.196 Historia natural de Macrotera pipiyolin (Hymenoptera: Andrenidae) en la Estación de Biología Chamela, Jalisco, México Natural history of Macrotera pipiyolin (Hymenoptera: Andrenidae) in the Estación de Biología Chamela, Jalisco, Mexico Beatriz Rodríguez-Velez1* y Ricardo Ayala2 1Instituto de Biología, Departamento de Zoología, Universidad Nacional Autónoma de México. Apartado postal 70-153, 04510 México, D.F., México. 2Estación de Biología Chamela, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado postal 21, San Patricio, 48980 Jalisco, México. *Correspondencia: [email protected] Resumen. Se presenta información sobre la historia natural de Macrotera pipiyolin; el estudio se realizó en la Estación de Biología Chamela, Instituto de Biología, UNAM, entre junio y agosto de 1994. Las abejas emergieron poco después del inicio de la temporada de lluvias, su pico de actividad fue a principio de julio, presentando alrededor de 35 días activos. La actividad concuerda con la fl oración de Opuntia excelsa, planta de la que obtiene sus recursos. Las cópulas ocurren sobre las fl ores y los machos muestran 3 tipos de estrategias reproductivas: macho dominante, subordinado y patrullero. Las hembras son poliándricas y los machos polígamos. Las hembras construyen los nidos generalmente cerca de O. excelsa y tienen una distribución gregaria. Los nidos son simples, con un túnel principal y celdas, las que se cierran y desconectan del tunel una vez que se han aprovisionado y en ellas se ha depositado un huevo. Para la provisión de una celda se requiere de 2 a 18 cargas de polen. -
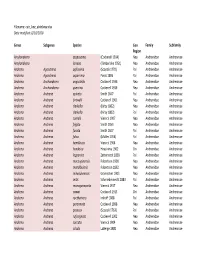
Anthophila List
Filename: cuic_bee_database. -

Components of Nest Provisioning Behavior in Solitary Bees (Hymenoptera: Apoidea)*
Apidologie 39 (2008) 30–45 Available online at: c INRA/DIB-AGIB/ EDP Sciences, 2008 www.apidologie.org DOI: 10.1051/apido:2007055 Review article Components of nest provisioning behavior in solitary bees (Hymenoptera: Apoidea)* John L. Neff Central Texas Melittological Institute, 7307 Running Rope, Austin, Texas 78731, USA Received 13 June 2007 – Revised 28 September 2007 – Accepted 1 October 2007 Abstract – The components of nest provisioning behavior (resources per cell, transport capacity, trip dura- tion, trips per cell) are examined for a data set derived from the literature and various unpublished studies. While there is a trade-off between transport capacity and trips required per cell, the highest provisioning rates are achieved by bees carrying very large pollen loads at intermediate trip durations. Most solitary bees appear to be either egg or resource limited, so sustained provisioning rates over one cell per day are unusual. Provisioning rate / body size / transport capacity / solitary bees / trip duration 1. INTRODUCTION tradeoffs between provisioning one large cell or two small cells direct, indirect, or nonex- There is a vast literature on the foraging tac- istent? Are provisioning rates related to body tics of bees: what kinds of flowers to visit, how size? long to spend on a flower, how many flowers Provisioning rate is a general term and can in an inflorescence to visit, when to turn, and simply refer to the rate (mass per unit time) so forth. Much of this work is done within a at which foragers bring various food items framework of whether a forager should max- (pollen, nectar, oils, carrion, or whatever) to imize harvesting rate or foraging efficiency their nests. -

O'donnell, S. 1994. Nestmate Copulation in the Neotropical Eusocial Wasp Polistes Instabilis De Saussure
NESTMATE COPULATION IN THE NEOTROPICAL EUSOCIAL WASP POLISTES INSTABILIS DE SAUSSURE (HYMENOPTERA: VESPIDAE) BY SEAN O'DoNNELL Department of Entomology University of California Davis, CA 95616 ABSTRACT Copulation between individually marked nestmates of the Neotropical eusocial wasp Polistes instabilis is reported. Polistes copulations are rarely observed in the field; this observation is the first documented case of nestmate copulation in the genus. Hamil ton ( 1964) predicted high rates of nestmate copulation for tropical Polistes populations which have asynchronous colony cycles rela tive to temperate populations. Nestmate copulation may lead to rel atively high levels of inbreeding in tropical Polistes species. INTRODUCTION Relative to temperate congeners, populations of tropical Polistes species are asynchronous in colony foundation and repro duction (West Eberhard 1969; Richards 1978; Young 1986). Even in seasonal tropical habitats, Polistes colonies can be found simul taneously in different stages of development (S. O'D. pers. obs.). Hamilton (1964) predicted that copulation among nestmates will be most common in tropical Polistes species because when repro ductives emerge asynchronously from different colonies, the avail ability of non-nestmate mating partners is reduced. Polistes nestmates tend to be close genetic relatives (reviewed in Reeve 1991 ); therefore, rates of inbreeding should be higher in tropical than in temperate Polistes species if nestmate copulations are fre quent. These predictions remain largely untested. Here I report copulation between individually marked nestmates of the Neotropical eusocial wasp Polistes instabilis. Polistes copu lations are rarely observed in the field (West Eberhard 1969; Manuscript received 21 December 1993. 33 34 Psyche [Vol. 101 1994] O'Donnell 35 Noonan 1978; Hook 1982), and copulation between identified nest forager, ranking fourth highest among 22 workers in rate of nectar mates has not previously been reported. -

Proceedings of the Tenth Forum Herbulot 2018. Integrative Taxonomy, a Multidisciplinary Approach to Answer Compli- Cated Taxonomic Questions
SPIXIANA 42 2 291-320 München, Dezember 2019 ISSN 0341-8391 Proceedings of the tenth FORUM HERBULOT 2018. Integrative taxonomy, a multidisciplinary approach to answer compli- cated taxonomic questions (Stuttgart, Germany, 11-16 June 2018) Axel Hausmann & Hossein Rajaei (eds) Hausmann, A. & Rajaei, H. (eds) 2019. Proceedings of the tenth FORUM HERBULOT 2018. Integrative taxonomy, a multidisciplinary approach to answer complicated taxonomic questions (Stuttgart, Germany, 11-16 June 2018). Spixiana 42 (2): 291- 320. The tenth International Congress of FORUM HERBULOT on “Integrative taxonomy, a multidisciplinary approach to answer complicated taxonomic questions” took place in the Staatliches Museum für Naturkunde Stuttgart (SMNS), from 11.- 16.06.2018, with 77 participants and 52 scientific presentations. The proceedings provide short information on the meeting and the abstracts of the oral presenta- tions. Axel Hausmann (corresponding author), SNSB – ZSM, Bavarian State Collection of Zoology, Münchhausenstr. 21, 81247 Munich, Germany; e-mail: [email protected] Short report and results Axel Hausmann & Hossein Rajaei Hausmann, A. & Rajaei, H. 2019. Short report and results. Pp. 291-292 in: Hausmann, A. & Rajaei, H. (eds). Proceedings of the tenth FORUM HERBULOT 2018. Integrative taxonomy, a multidisciplinary approach to answer complicated taxonomic questions (Stuttgart, Germany, 11-16 June 2018). Spixiana 42 (2). Axel Hausmann (corresponding author), SNSB – ZSM, Bavarian State Collection of Zoology, Münchhausenstr. 21, 81247 Mu- nich, Germany; e-mail: [email protected] The meeting was organized by an organization The conference started with a lecture on the ground- team of the ‘Staatliches Museum für Naturkunde breaking effects of “Willi Hennig and the synthesis of Stuttgart’ (SMNS).