Boreale Lauvskoger I Norge
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

4634 72Dpi.Pdf (6.278Mb)
Norsk institutt for vannforskning RAPPORT Hovedkontor Sørlandsavdelingen Østlandsavdelingen Vestlandsavdelingen Akvaplan-niva Postboks 173, Kjelsås Televeien 3 Sandvikaveien 41 Nordnesboder 5 0411 Oslo 4879 Grimstad 2312 Ottestad 5005 Bergen 9296 Tromsø Telefon (47) 22 18 51 00 Telefon (47) 37 29 50 55 Telefon (47) 62 57 64 00 Telefon (47) 55 30 22 50 Telefon (47) 77 75 03 00 Telefax (47) 22 18 52 00 Telefax (47) 37 04 45 13 Telefax (47) 62 57 66 53 Telefax (47) 55 30 22 51 Telefax (47) 77 75 03 01 Internet: www.niva.no Tittel Løpenr. (for bestilling) Dato Foreløpig forslag til system for typifisering av norske 4634-2003 10.02.03 ferskvannsforekomster og for beskrivelse av referansetilstand, samt forslag til referansenettverk Prosjektnr. Undernr. Sider Pris 21250 93 Forfatter(e) Fagområde Distribusjon Anne Lyche Solheim, Tom Andersen, Pål Brettum, Lars Erikstad (NINA), Arne Fjellheim (LFI, Stavanger museum), Gunnar Halvorsen (NINA), Trygve Hesthagen (NINA), Eli-Anne Lindstrøm, Geografisk område Trykket Marit Mjelde, Gunnar Raddum (LFI, Univ. i Bergen), Tuomo Norge NIVA Saloranta, Ann-Kristin Schartau (NINA), Torulv Tjomsland og Bjørn Walseng (NINA) Oppdragsgiver(e) Oppdragsreferanse SFT, DN, NVE Sammendrag Innføringen av Rammedirektivet for vann (”Vanndirektivet”) medfører at Norges vannforekomster innen utgangen av 2004 skal inndeles og beskrives etter gitte kriterier. Et av kriteriene er en typeinndeling etter fysiske og kjemiske faktorer. Denne typeinndelingen danner grunnlaget for overvåkning og bestemmelse av økologisk referansetilstand for påvirkede vannforekomster. Vanndirektivet gir valg mellom å bruke en predefinert all-europeisk typologi (”System A”), eller å etablere en nasjonal typologi som forutsettes å gi bedre og mer relevant beskrivelse enn den all-europeiske, og som må inneholde visse obligatoriske elementer (”System B”). -

Notat 30.11.2020, Forslag Til Tiltaksorientert Overvåkingsprogram
NOTAT 30. november 2020 Mottakere: Bergmesteren Raudsand AS ved Keith Roebuck. Utarbeidet av NIVA v/: Merete Schøyen. Kopi: NIVAs arkiv. Journalnummer: 0462/20. Prosjektnummer: O-190224. Forslag til tiltaksorientert overvåkingsprogram for Bergmesteren Raudsand AS i henhold til vannforskriften NIVA har fått i oppdrag å lage et overvåkingsprogram for Bergmesteren Raudsand AS, for å overvåke kjemisk og økologisk tilstand i vannforekomst Tingvollfjorden ved Raudsand. Det vises til telefonsamtale med Bergmesteren Raudsand AS ved Keith Roebuck og Norsk institutt for vannforskning (NIVA) ved Marijana Stenrud Brkljacic den 12.10.2020. Bergmesteren Raudsand AS skal levere overvåkingsprogram til Miljødirektoratet jamfør «Tillatelse etter forurensningsloven for Deponi 2 og Møllestøvdeponiet» datert 29.08.2019. NIVA har utarbeidet et forslag til overvåkingsprogram basert på funn og anbefalinger ved forrige miljøovervåking i 2019 (Brkljacic m fl. 2020) og krav spesifisert i tillatelsen med spesielt fokus på kapittel 3.9 (Overvåking). Hensikten med overvåkingen er å identifisere hvorvidt bedriftens utslipp påvirker vannforekomstens økologiske og kjemiske tilstand. Bedriften ønsker at programforslaget skal inkludere et forslag til frekvens. Miljødirektoratet vil vurdere behovet for videre overvåking og eventuell frekvens etter at resultatet fra første runde av overvåkingen foreligger. 1 Beskrivelse av bedriften og utslippet 1.1 Kort beskrivelse av virksomheten Bergmesteren Raudsand AS ligger i Tingvollfjorden i Molde kommune i Møre og Romsdal (Figur 1). Bedriften tilhører sektoren deponier og bransjen «Behandling og disponering av farlig avfall». _________________________________________________________________________ 1 Bergmesteren Raudsand AS Figur 1. Bergmesteren Raudsand AS ligger i Tingvollfjorden i Molde kommune i Møre og Romsdal (hentet fra Øxnevad 2015). Bedriften har deponier for produksjonsavfall fra lokale bedrifter som har drevet sekundær og tertiær aluminiumsproduksjon. -

Namn Og Nemne 28 – 2011
namn Manuskript til Namn og Nemne Redaksjonen vil helst ha manus elektronisk som vedlegg til e-post. Blir det og brukt spesialteikn, ønskjer vi også ei pdf-fil av artikkelen. nem Artiklar på skandinaviske språk skal ha eit samandrag på engelsk eller tysk. Samandraget skal kort gje innsyn i dei viktigaste momenta i artikkelen. n Sitat som er lengre enn tre maskinskrivne liner, skal skiljast frå teksten ved e 28 – 2011 innrykking. Fotnotar (merknader) bør brukast så lite som råd; dei bør oftast takast namn inn i teksten. Eventuelle fotnotar skal samlast i slutten av artikkelen. Nummertilvisingane i teksten skal ikkje ha parentes kring seg, og dei skal og nemne plasserast etter skiljeteikn. Litteraturtilvisingane skal om mogleg gjerast ved forfattarnamn og Tidsskrift for utgjevingsår. Døme: ... (Indrebø 1924: 98), eller: Gustav Indrebø (1924: 98) norsk meiner ... , eller: Som opplyst i Indrebø 1924: 98, ... Arbeid som er fotografisk oppattprenta, skal siterast etter originalutgava (av forskingshistoriske grunnar), namnegransking altså ikkje Olsen, M. 1978. Torp, A. 1963, Aasen, I. 1918 eller 1977. Allment kjende tidsskrift bør ha forkorta titlar (t.d. ANF, APhS, MM, NoB, SvLm); det same kan gjelde kjende seriar (t.d. DS [Danmarks Stednavne], NG [Norske Gaardnavne], men då med tilvising til forfattar eller utgjevar. Undertitlar og serieopplysningar trengst til vanleg ikkje. Døme: Andersson, T. 1983: Personnamn. Till begreppets avgränsning. NORNA-rapporter 23. 9–23. Hovda, P. 1966: Norske elvenamn. Oslo–Bergen. [Hovda, P.] 1978: Frå hav til hei. Oslo–Bergen–Tromsø. Indrebø, G. (red.) 1920: Sverris Saga etter Cod. AM 327 4°. Kristiania: Den Norske Historiske Kildeskriftkommission. Indrebø, G. 1921: Stadnamni i ei fjellbygd. -

Stabilitet Langs Namsen Utbedring Av Gamle Sikringstiltak Mot Ras Og Erosjon Anders Bjordal Mads Johnsen
Norges vassdrags- og energidirektorat Telefon: 22 95 95 95 Middelthunsgate 29 Telefaks: 22 95 90 00 Postboks 5091 Majorstua Internett: www.nve.no 0301 Oslo Stabilitet langs Namsen Utbedring av gamle sikringstiltak mot ras og erosjon Anders Bjordal Mads Johnsen 5 2007 RAPPORT Stabilitet langs Namsen Utbedring av gamle sikringstiltak mot ras og erosjon Norges vassdrags- og energidirektorat 2007 Rapport nr 5 Utbedring av gamle sikringstiltak mot ras og erosjon Utgitt av: Norges vassdrags- og energidirektorat Forfatter: Anders Bjordal, Mads Johnsen Medforfatter: Geir B Hagen, Barbro Folde, Tharan Fergus Trykk: NVEs hustrykkeri Opplag: 50 Forsidefoto: Flyfoto av Namsen ved Vibstad, anleggsarbeid på 70-tallet og anleggsarbeid i dag. Foto: NVEs fotodatabase ISBN: 978-82-410-0630-2 ISSN: 1501-2832 Sammendrag: Rapporten beskriver alle sikringsanlegg langs Namsen fra Namsos til Grong. Bakgrunnen til alle anleggene er beskrevet og alle sikringsanleggene er vist på oversiktskart. De anleggene som har behov for vedlikehold, reparasjon eller miljømessig oppgradering er beskrevet nærmere. For disse anleggene er det laget tiltaksbeskrivelse, detaljerte kart og kostnadsoverslag. Emneord: Flom, erosjon, sikringstiltak, erosjonssikringstiltak, miljøtiltak kvikkleire, kvikkleireras. Sammendrag I all tid har det gått store ras langs Namsen. Etter store skred i Namsen i 1959 og registrering av erosjonstilstanden i vassdraget i 1961, ble det i 1960 og 70-årene gjennomført et storstilt forbyggingsprogram i Namsen for å sikre mot videre erosjon og utrasing. I det store og dype elvefaret tillot ikke utstyr og arbeidsmetoder den gang en tilstrekkelige god utførelse av forbygningene. Det ble heller ikke lagt særlig vekt på istandsetting og tilrettelegging av adkomstvegen, landskap og miljø. I dag er et stort antall av de gamle forbygningene skadet og trenger reparasjon og miljømessig opprusting. -

St.Prp. Nr. 75 (2003–2004)
Utenriksdepartementet St.prp. nr. 75 (2003–2004) Supplering av Verneplan for vassdrag Innhold Del I Den generelle del . 7 8.3 Den samlede Verneplan for vassdrag . 28 1 Innledning og sammendrag . 9 9 De enkelte objekter . 31 2 Verneplan for vassdrag . 10 9.1 Hedmark . 31 9.1.1 Imsa . 31 3 Bakgrunnen for 9.1.2 Sølna . 32 verneplansuppleringen 9.1.3 Tunna . 32 og organiseringen av arbeidet . 11 9.2 Oppland. 33 3.1 Bakgrunnen for 9.2.1 Vismunda . 33 verneplansuppleringen . 11 9.2.2 Vassdragene i Øvre Otta . 34 3.2 Organiseringen av arbeidet. 11 9.2.2.1 Tora . 34 9.2.2.2 Glitra . 34 4 Utvelgelsen av vassdrag for 9.2.2.3 Mosagrovi . 35 vurdering . 12 9.2.2.4 Måråi . 35 4.1 Grunnlaget for utvelgelsen av 9.2.2.5 Åfåtgrovi . 35 vassdrag for vurdering. 12 9.2.3 Jora . 36 4.2 Innkomne forslag til vurdering 9.2.4 Vinda . 37 for vern . 12 9.3 Buskerud . 38 4.3 Forslagene fra styringsgruppen 9.3.1 Nedalselva . 38 og NVE . 12 9.3.2 Dagali (Godfarfoss) . 39 4.4 Departementshøringen . 13 9.4 Vestfold . 40 9.4.1 Dalelva . 40 5 Forholdet til Samlet plan og 9.5 Telemark. 41 nasjonale laksevassdrag . 18 9.5.1 Kåla . 41 5.1 Forholdet til Samlet plan . 18 9.5.2 Digeråi . 41 5.2 Forholdet til andre runde av 9.5.3 Rauda . 42 nasjonale laksevassdrag . 18 9.5.4 Skoevassdraget . 43 9.6 Aust-Agder . 44 6 Verneplanens virkninger. 20 9.6.1 Tovdalsvassdraget ovenfor 6.1 Verneplanens rettslige status . -
3323 72Dpi.Pdf (329.1Kb)
1 Norsk institutt for vannforskning O-91050 Landsomfattende trofiundersøkelse av innsjøer Problemnotat om tilfeldig utvalg av innsjøer 1 FORORD Bakgrunnen for dette notatet var diskusjoner i SFT og NIVA høsten 1994 om behovet for at innsjøer i landsomfattende undersøkelser skal trekkes ut statistisk tilfeldig for å tilfredsstille de aktuelle målsetninger med undersøkelsene. Diskusjonene har gått parallelt for "Landsomfattende trofiundersøkelse av norske innsjøer" og "1000-sjøer undersøkelsen av forsuring". Sistnevnte skal gjennomføres på nytt i 1995, og det er planer om å utvide antallet innsjøer som skal undersøkes. Da målsettingen med de to undersøkelsene er noe forskjellig - og ikke minst fordi de fenomenene en skulle studere var ulikt fordelt over landet, ble det også diskutert om strategien for utvalg av innsjøer kan/bør være forskjellig. For "trofiundersøkelsen" ble det avholdt et diskusjonsmøte i SFT den 18. januar 1995. Møtet konkluderte med at det er hensiktsmessig å fortsette undersøkelsen med det utvalget av innsjøer som ble gjort i 1988, med enkelte tillegg i 1992. Det var også enighet om behovet for å utarbeide et notat med presentasjon av endel synspunkter på tilfeldig utvalg av innsjøer. Synspunktene representerer primært de sider av problematikken som er relevante for trofiundersøkelsen, og er ikke nødvendigvis dekkende for andre undersøkelser. Gunnar Severinsen har tilrettelagt data fra Vassdragsregisteret og bidratt ved bearbeidingen av disse. Oslo 31. mai 1995 Bjørn Faafeng 2 INNHOLD side FORORD 1 INNHOLD 2 1. KONKLUSJONER 3 2. TILFELDIG UTVALG 4 2.1 Definisjon og utvalg 4 2.2 Stratifisert tilfeldig utvalg 4 2.3 Tilfeldig utvalg eller ikke? - målsetting og rammebetingelser avgjør! 5 3. -

Verneplan for Ursvatnet Etterseterbekken Oksbåsen Og
+ Verneplan for skog – frivillig vern Høring av forslag om vern av fire områder i tre kommuner i Nordland Miljøvernavdelinga Rapportnummer/år Foto: Forsidebilde: Ingvild Gabrielsen/Fylkesmannen i Nordland 1 Navn på rapport Forord Innholdsfortegnelse Oppsummering .................................................................................... 4 1 Innledning ...................................................................................... 4 2 Lovgrunnlag .................................................................................... 5 3 Verneplanposessen ........................................................................ 6 3.1 Generelt ............................................................................................................................................ 6 3.2 Naturfaglige registreringer ............................................................................................................. 7 3.3 Oppstart av verneplanarbeidet ..................................................................................................... 7 3.4 Saksbehandling ................................................................................................................................ 9 3.5 Verneplanens omfang ................................................................................................................... 10 3.6 Konsekvenser av verneforslagene ............................................................................................... 11 3.6.1 Skogbruk .................................................................................................................................... -
Jordarter V E U N O T N a Leirpollen
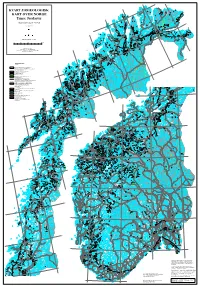
30°E 71°N 28°E Austhavet Berlevåg Bearalváhki 26°E Mehamn Nordkinnhalvøya KVARTÆRGEOLOGISK Båtsfjord Vardø D T a e n a Kjøllefjord a n f u j o v r u d o e Oksevatnet t n n KART OVER NORGE a Store L a Buevatnet k Geatnjajávri L s Varangerhalvøya á e Várnjárga f g j e o 24°E Honningsvåg r s d Tema: Jordarter v e u n o t n a Leirpollen Deanodat Vestertana Quaternary map of Norway Havøysund 70°N en rd 3. opplag 2013 fjo r D e a T g tn e n o a ra u a a v n V at t j j a n r á u V Porsanger- Vadsø Vestre Kjæsvatnet Jakobselv halvøya o n Keaisajávri Geassájávri o Store 71°N u Bordejávrrit v Måsvatn n n i e g Havvannet d n r evsbotn R a o j s f r r Kjø- o Bugøy- e fjorden g P fjorden 22°E n a Garsjøen Suolo- s r Kirkenes jávri o Mohkkejávri P Sandøy- Hammerfest Hesseng fjorden Rypefjord t Bjørnevatn e d n Målestokk (Scale) 1:1 mill. u Repparfjorden s y ø r ø S 0 25 50 100 Km Sørøya Sør-Varanger Sállan Skáiddejávri Store Porsanger Sametti Hasvik Leaktojávri Kartet inngår også i B áhèeveai- NASJONALATLAS FOR NORGE 20°E Leavdnja johka u a Lopphavet -

Sunndal and Tingvoll
2008-09 Lokalguide Sunndal and Tingvoll Toppen av Fjord Norge Spektakulære Aursjøvegen Lokale spesialiteter Kulturkalender Se side 7 Se side 17 Se side 26 R uide okalg l & Rin L urnada Halsa, S ide & algu verøy Lok nd, A ansu Kristi Lok R alguide Smøla & Aure L oka Su lgu nnd ide al & Ting voll Mardalsfossen Innhold/Content/Inhalt Velkommen til Tingvoll og Sunndal! Sunndal s 4 Tingvoll s 5 Turistinformasjonen i Sunndal På toppen av Fjord Norge ligger et kontrastfylt landskap. Norges vakreste fjelldal Innerda - Welcome! Wilkommen ! Innerdalen s 6 Auragata 3 len, den spektakulære Aursjøvegen, mektige Trollheimen og andre flotte vandreområder. Aursjøvegen s 7 6600 Sunndalsøra Gode lakseelver, spennende havfiske, rorbuferie, levende kystsamfunn, sterke trebåttradi - Explore the most beautiful valley of Nor - Die Landschaft im nördlichen Fjordnorwe - Tel: (+47) 71 68 99 70 sjoner, klippfisk, opera og Atlanterhavsveien – opplevelsene venter deg. Velkommen til way, Innerdalen. Drive the spectacular gen ist äußerst kontrastreich: Innerdalen, Seterlandet og kulturminner s 8 Fax: (+47) 71 68 99 71 Nordmøre! mountain road of Aursjøvegen. Enjoy the das schönste Bergtal Norwegens, der Fiske s 9 E-post: [email protected] mighty mountain range of Trollheimen, spektakuläre Aursjøvegen, das mächtige Fantastiske fosser s 10 I Tingvoll finner du mange spennende produsenter av lokalmat. Sunndal har en storslått hike in scenic surroundings. Salmon rivers Trollheimen und andere herrliche Wan - Turistinformasjonen i Tingvoll fjellverden. Gå ikke glipp av Norges vakreste fjelldal, Innerdalen, som ligger midt mellom and deep sea fishing, rorbu holidays, li - dergebiete. Lachsflüsse, Fischen auf Øksendalsøra s 11 c/o Tingvoll Fjordhotell Tingvoll og Sunndal. -

Regional Plan for Vannforvaltning I Vannregion Nordland Og Jan Mayen (2016-2021)
Regional plan for vannforvaltning i vannregion Nordland og Jan Mayen (2016-2021) 1 Hjemmeside: www.vannportalen.no/nordland Vedtatt av Nordland fylkesting dato og saksnummer. Vannregionmyndigheten i Nordland Nordland fylkeskommune Fylkeshuset 8048 BODØ Epost: [email protected] 2 Forord 3 Sammendrag Hovedmålsettingen med Regional plan for vannforvaltning i vannregion Nordland og Jan Mayen er å gi en enkel og oversiktlig framstilling av hvordan vannmiljøet og vannressursene i vannregionen skal forvaltes i et langsiktig perspektiv. Planen er utarbeidet i henhold til forskrift om rammer for vannforvaltningen (vannforskriften) og plan og bygningsloven (pbl) kap 8. Planen skal rulleres hvert 6. år og danner grunnlag for en systematisk og helhetlig vannforvaltning. Planen setter miljømål for alle vannforekomstene i fylket. Målene vil være styrende for videre forvaltning. Den geografiske avgrensingen er Nordland fylke med justeringer mot Troms og Trøndelag da vannregionen er definert av nedbørsfelt og ikke administrative grenser. I tillegg er Nordland en internasjonal vannregion. Det innebærer at deler vannregioner på svensk side inngår i planarbeidet. Fylkestinget i Nordland skal imidlertid kun fatte vedtak om det vannet som er innen norsk territorium. Jan Mayen inngår også i vannregionen. Dette er norsk territorium, men uten fastboende gjelder ikke plan- og bygningsloven så fylkestinget har ingen vedtaksmyndighet her. Forvaltningsplanen gjelder for 2016-2021 bygger på en rullering av Forvaltningsplan for vannregion Nordland (2010-2015) som ble vedtatt av Fylkestinget i Nordland i 2009. Denne planen omhandler kun vannområde Ranfjorden og inngikk i en nasjonal frivillig pilotfase. Planfasen fra 2016-2021 er den første Norge har forpliktet seg til gjennom EØS-avtalen. Avtalen bygger på EUs vanndirektiv som gjennomføres i hele Europa etter samme tidsskjema. -

Samordning Av Lokaliteter Og Framtidige Utfordringer
Statlig program for forurensingsovervåking Nasjonale programmer for innsjøovervåking Samordning av lokaliteter og framtidige utfordringer TA-1949/2003 ISBN 82-577-4320-8 Referer til denne rapporten som: SFT, 2003. Nasjonale programmer for innsjøovervåking - Samordning av lokaliteter og framtidige utfordringer. (TA-1949/2003) Oppdragsgivere: Statens forurensningstilsyn Postboks 8100 Dep. 0032 Oslo Utførende institusjoner: Norsk institutt for naturforskning Tungasletta 2 7485 Trondheim Norsk institutt for vannforskning Postboks 173 Kjelsås 0411 Oslo Akvaplan-niva AS 9296 Tromsø Forord SFT har i e-mail 24.06.2002 anmodet NIVA om å koordinere et prosjektforslag i samarbeid med Akvaplan-niva og NINA angående fremtidig overvåking av innsjøer og koordinering av ulike overvåkingsprogrammer. Det er et ønske fra SFT om å samordne utvalget av innsjølokaliteter i seks nasjonale overvåkingsprogrammer og se dette i lys av framtidige planer for overvåking og i forbindelse med implementering av Vanndirektivet. Målsetningen er å skaffe en bedre oversikt over de aktivitetene som har foregått til nå, for om mulig å redusere kostnader ved fremtidig feltarbeid og innsamling. Videre ønsker SFT å dra nytte av informasjonen fra forskjellige overvåkingsprogrammer for å få større kunnskap om tilstanden i hver enkelt innsjø. Denne oversikten skal også være utgangspunkt for utvalg av lokaliteter til framtidig overvåking. Denne rapporten gir en oversikt over lokaliteter og aktiviteter i ca. 1000 lokaliteter i 6 nasjonale overvåkingsprogrammer og diskuterer mulighetene -

3. Rødlisteartar I Møre Og Romsdal – Planter, Sopp Og
1 FORORD (Frå Fylkesmannen) Denne rapporten er eit nytt bidrag i arbeidet med å få oversikt over dei biologiske verdiane i Møre og Romsdal, og såleis ein av fleire måtar å følgje opp den nasjonale miljøvernpolitikken, "føre-var"-prinsippet samt anna internasjonalt avtaleverk. I St. meld. nr. 58 om miljøvernpolitikk for ei berekraftig utvikling blir det mellom anna opplyst at tap av biologisk mangfald er eit alvorleg globalt problem som er i ferd med å undergrave grunnlaget for ei berekraftig utvikling. Denne rapporten viser at det også i Møre og Romsdal er behov for ei bevisst haldning til farane mennesket sine daglege inngrep i naturen kan føre til. Fylkesmannen ser det i ein slik samanheng som sers viktig at forskjellige sektorar samt kommunane har ein best mogleg kjennskap til artsmangfaldet i fylket. Kunnskap om det biologiske mangfaldet er avgjerande for at til dømes kommunale planleggarar, landbruksnæringa og andre som arbeider med planlegging og gjennomføring av tiltak rundt i fylket kan ta dei nødvendige miljøomsyn i sitt arbeid. Vi håpar at rapporten også kan auke den almenne interessa og forståinga av naturverdiane i fylket. Fylkesmannen har finansiert utarbeiding av rapporten, medan innsamla data og vurderingar står for forfattarane si rekning. Molde 02.11.1998 Per Fredrik Brun Fylkesmiljøvernsjef (Fra forfatterne) En rekke personer har bidratt til denne rapporten. Siden en betydelig del av materialet (57%) er egne funn skylder vi alle som har hjulpet oss i bestemmelsesarbeidet en stor takk. En del av disse er allerede takket i tidligere rapporter hvor en stor del av materialet er presentert. Likevel er det en god del funn i den foreliggende rapporten som ikke har stått på trykk tidligere.