Types of Skeletal Systems
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Applications of Chondrocyte-Based Cartilage Engineering: an Overview
Hindawi Publishing Corporation BioMed Research International Volume 2016, Article ID 1879837, 17 pages http://dx.doi.org/10.1155/2016/1879837 Review Article Applications of Chondrocyte-Based Cartilage Engineering: An Overview Abdul-Rehman Phull,1 Seong-Hui Eo,1 Qamar Abbas,1 Madiha Ahmed,2 and Song Ja Kim1 1 Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongjudaehakro 56, Gongju 32588, Republic of Korea 2Department of Pharmacy, Quaid-i-Azam University, Islamabad 45320, Pakistan Correspondence should be addressed to Song Ja Kim; [email protected] Received 14 May 2016; Revised 24 June 2016; Accepted 26 June 2016 Academic Editor: Magali Cucchiarini Copyright © 2016 Abdul-Rehman Phull et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Chondrocytes are the exclusive cells residing in cartilage and maintain the functionality of cartilage tissue. Series of biocomponents such as different growth factors, cytokines, and transcriptional factors regulate the mesenchymal stem cells (MSCs) differentiation to chondrocytes. The number of chondrocytes and dedifferentiation are the key limitations in subsequent clinical application of the chondrocytes. Different culture methods are being developed to overcome such issues. Using tissue engineering and cell based approaches, chondrocytes offer prominent therapeutic option specifically in orthopedics for cartilage repair and to treat ailments such as tracheal defects, facial reconstruction, and urinary incontinence. Matrix-assisted autologous chondrocyte transplantation/implantation is an improved version of traditional autologous chondrocyte transplantation (ACT) method. An increasing number of studies show the clinical significance of this technique for the chondral lesions treatment. -

Pg 131 Chondroblast -> Chondrocyte (Lacunae) Firm Ground Substance
Figure 4.8g Connective tissues. Chondroblast ‐> Chondrocyte (Lacunae) Firm ground substance (chondroitin sulfate and water) Collagenous and elastic fibers (g) Cartilage: hyaline No BV or nerves Description: Amorphous but firm Perichondrium (dense irregular) matrix; collagen fibers form an imperceptible network; chondroblasts produce the matrix and when mature (chondrocytes) lie in lacunae. Function: Supports and reinforces; has resilient cushioning properties; resists compressive stress. Location: Forms most of the embryonic skeleton; covers the ends Chondrocyte of long bones in joint cavities; forms in lacuna costal cartilages of the ribs; cartilages of the nose, trachea, and larynx. Matrix Costal Photomicrograph: Hyaline cartilage from the cartilages trachea (750x). Thickness? Metabolism? Copyright © 2010 Pearson Education, Inc. Pg 131 Figure 6.1 The bones and cartilages of the human skeleton. Epiglottis Support Thyroid Larynx Smooth Cartilage in Cartilages in cartilage external ear nose surface Cricoid Trachea Articular Lung Cushions cartilage Cartilage of a joint Cartilage in Costal Intervertebral cartilage disc Respiratory tube cartilages in neck and thorax Pubic Bones of skeleton symphysis Meniscus (padlike Axial skeleton cartilage in Appendicular skeleton knee joint) Cartilages Articular cartilage of a joint Hyaline cartilages Elastic cartilages Fibrocartilages Pg 174 Copyright © 2010 Pearson Education, Inc. Figure 4.8g Connective tissues. (g) Cartilage: hyaline Description: Amorphous but firm matrix; collagen fibers form an imperceptible network; chondroblasts produce the matrix and when mature (chondrocytes) lie in lacunae. Function: Supports and reinforces; has resilient cushioning properties; resists compressive stress. Location: Forms most of the embryonic skeleton; covers the ends Chondrocyte of long bones in joint cavities; forms in lacuna costal cartilages of the ribs; cartilages of the nose, trachea, and larynx. -

GLOSSARY of MEDICAL and ANATOMICAL TERMS
GLOSSARY of MEDICAL and ANATOMICAL TERMS Abbreviations: • A. Arabic • abb. = abbreviation • c. circa = about • F. French • adj. adjective • G. Greek • Ge. German • cf. compare • L. Latin • dim. = diminutive • OF. Old French • ( ) plural form in brackets A-band abb. of anisotropic band G. anisos = unequal + tropos = turning; meaning having not equal properties in every direction; transverse bands in living skeletal muscle which rotate the plane of polarised light, cf. I-band. Abbé, Ernst. 1840-1905. German physicist; mathematical analysis of optics as a basis for constructing better microscopes; devised oil immersion lens; Abbé condenser. absorption L. absorbere = to suck up. acervulus L. = sand, gritty; brain sand (cf. psammoma body). acetylcholine an ester of choline found in many tissue, synapses & neuromuscular junctions, where it is a neural transmitter. acetylcholinesterase enzyme at motor end-plate responsible for rapid destruction of acetylcholine, a neurotransmitter. acidophilic adj. L. acidus = sour + G. philein = to love; affinity for an acidic dye, such as eosin staining cytoplasmic proteins. acinus (-i) L. = a juicy berry, a grape; applied to small, rounded terminal secretory units of compound exocrine glands that have a small lumen (adj. acinar). acrosome G. akron = extremity + soma = body; head of spermatozoon. actin polymer protein filament found in the intracellular cytoskeleton, particularly in the thin (I-) bands of striated muscle. adenohypophysis G. ade = an acorn + hypophyses = an undergrowth; anterior lobe of hypophysis (cf. pituitary). adenoid G. " + -oeides = in form of; in the form of a gland, glandular; the pharyngeal tonsil. adipocyte L. adeps = fat (of an animal) + G. kytos = a container; cells responsible for storage and metabolism of lipids, found in white fat and brown fat. -

Bone Markings / Features on Bones
08/05/2016 Bone Markings : Skeletal System Search Custom Search Like Tweet Home Health News Human Body Biology Chemistry Glossary Textbooks Bone Disorders Ads by Google ► Bone Tissue ► Bone Marrow ► Human Skull Bone ► Bone on Bone Knee Sun 8 May 2016 Bone Markings / Features on Bones Human Body Study Section Bone markings and the features of bones (including the correct words used to describe them) are often required by firstlevel courses in human anatomy and associated health science subjects. It is important to be familiar with the terminology used to Human Body Index refer to bone markings in order to communicate effectively with professionals involved in healthcare, research, forensics, and Health Glossary related disciplines. More about Bones and the Skeletal System: The following terms used to describe bone markings or features on bones are in alphabetical order with short definitions: Human Skeleton Axial and Appendicular Word / Term Meaning / Description Type of Example(s) Skeleton (Bone Marking or bone The Structure and Feature) marking Functions of Bones Types of Bones 1. Angle A corner Feature of Inferior angle (lower) and superior angle (upper) are Bone Markings & Features shape of bone the rounded angles or "corners" of the scapula. on Bones Disorders of the Skeletal 2. Body The main portion of a bone The diaphysis of long bones such as the humerus. System Curvature of the Spine 3. Condyle Rounded bump or large rounded Process The medial condyle of the femur (bone), upperleg. prominence. Such rounded surfaces forms joints Types of Joints usually fit into a fossa on another bone to Specific bones: form a joint. -

The Epiphyseal Plate: Physiology, Anatomy, and Trauma*
3 CE CREDITS CE Article The Epiphyseal Plate: Physiology, Anatomy, and Trauma* ❯❯ Dirsko J. F. von Pfeil, Abstract: This article reviews the development of long bones, the microanatomy and physiology Dr.med.vet, DVM, DACVS, of the growth plate, the closure times and contribution of different growth plates to overall growth, DECVS and the effect of, and prognosis for, traumatic injuries to the growth plate. Details on surgical Veterinary Specialists of Alaska Anchorage, Alaska treatment of growth plate fractures are beyond the scope of this article. ❯❯ Charles E. DeCamp, DVM, MS, DACVS athologic conditions affecting epi foramen. Growth factors and multipotent Michigan State University physeal (growth) plates in imma stem cells support the formation of neo ture animals may result in severe natal bone consisting of a central marrow P 2 orthopedic problems such as limb short cavity surrounded by a thin periosteum. ening, angular limb deformity, or joint The epiphysis is a secondary ossifica incongruity. Understanding growth plate tion center in the hyaline cartilage forming anatomy and physiology enables practic the joint surfaces at the proximal and distal At a Glance ing veterinarians to provide a prognosis ends of the bones. Secondary ossification Bone Formation and assess indications for surgery. Injured centers can appear in the fetus as early Page E1 animals should be closely observed dur as 28 days after conception1 (TABLE 1). Anatomy of the Growth ing the period of rapid growth. Growth of the epiphysis arises from two Plate areas: (1) the vascular reserve zone car Page E2 Bone Formation tilage, which is responsible for growth of Physiology of the Growth Bone is formed by transformation of con the epiphysis toward the joint, and (2) the Plate nective tissue (intramembranous ossifica epiphyseal plate, which is responsible for Page E4 tion) and replacement of a cartilaginous growth in bone length.3 The epiphyseal 1 Growth Plate Closure model (endochondral ossification). -

WHO Manual of Diagnostic Imaging Radiographic Anatomy and Interpretation of the Musculoskeletal System
The WHO manual of diagnostic imaging Radiographic Anatomy and Interpretation of the Musculoskeletal System Editors Harald Ostensen M.D. Holger Pettersson M.D. Authors A. Mark Davies M.D. Holger Pettersson M.D. In collaboration with F. Arredondo M.D., M.R. El Meligi M.D., R. Guenther M.D., G.K. Ikundu M.D., L. Leong M.D., P. Palmer M.D., P. Scally M.D. Published by the World Health Organization in collaboration with the International Society of Radiology WHO Library Cataloguing-in-Publication Data Davies, A. Mark Radiography of the musculoskeletal system / authors : A. Mark Davies, Holger Pettersson; in collaboration with F. Arredondo . [et al.] WHO manuals of diagnostic imaging / editors : Harald Ostensen, Holger Pettersson; vol. 2 Published by the World Health Organization in collaboration with the International Society of Radiology 1.Musculoskeletal system – radiography 2.Musculoskeletal diseases – radiography 3.Musculoskeletal abnormalities – radiography 4.Manuals I.Pettersson, Holger II.Arredondo, F. III.Series editor: Ostensen, Harald ISBN 92 4 154555 0 (NLM Classification: WE 141) The World Health Organization welcomes requests for permission to reproduce or translate its publications, in part or in full. Applications and enquiries should be addressed to the Office of Publications, World Health Organization, CH-1211 Geneva 27, Switzerland, which will be glad to provide the latest information on any changes made to the text, plans for new editions, and reprints and translations already available. © World Health Organization 2002 Publications of the World Health Organization enjoy copyright protection in accordance with the provisions of Protocol 2 of the Universal Copyright Convention. All rights reserved. -

Musculo-Skeletal System
Musculo-Skeletal System (Trunk, Limbs, and Head) somite: ectoderm dermatome General Statements: myotome Bilaterally, paraxial mesoderm become sclerotome neural crest somites and somitomeres. (Somitomeres develop ros- intermediate tral to the notochord in the head. They are like somites, but mesoderm neural tube smaller and less distinctly organized.) The mesoderm somatic mesoderm comprising each somite differentiates into three notochord regions: endoderm aorta — dermatome (lateral) which migrates to form dermis of the skin coelom — sclerotome (medial) forms most of the splanchnic mesoderm axial skeleton (vertebrae, ribs, and base of the skull). Mesoderm Regions — myotome (middle) forms skeletal mus- culature. Individual adult muscles are produced by merger of adjacent myotomes. Note: Nerves make early connections with adjacent myotomes and dermatomes, establishing a segmental innervation pattern. As myotome/dermatome cells migrate to assume adult positions, the segmental nerve supply must follow along to maintain its connection to the innervation target. (Recurrent laryngeal & phrenic nerves travel long distances because their targets migrated far away.) Skin. Consists of dermis and epidermis. Epidermis, including hair follicles & glands, is derived from ectoderm. Neural crest cells migrate into epidermis and become melanocytes. (Other neural crest cells become tactile disc receptors.) Dermis arises from mesoderm (dermatomes of somites). Each dermatome forms a continu- ous area of skin innervated by one spinal nerve. Because adjacent dermatomes overlap, a locus of adult skin is formed by 2 or 3 dermatomes, and innervated by 2 or 3 spinal nerves. Muscle. Muscles develop from mesoderm, except for muscles of the iris which arise from optic cup ectoderm. Cardiac and smooth muscles originate from splanchnic mesoderm. -
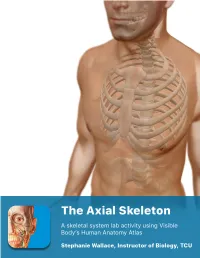
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

Download File
Cartilage Development and Maturation In Vitro and In Vivo Johnathan Ng Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Graduate School of Arts and Sciences Columbia University 2017 © 2017 Johnathan Ng All rights reserved Abstract Cartilage Development and Maturation In Vitro and In Vivo Johnathan Ng The articular cartilage has a limited capacity to regenerate. Cartilage lesions often result in degeneration, leading to osteoarthritis. Current treatments are mostly palliative and reparative, and fail to restore cartilage function in the long term due to the replacement of hyaline cartilage with fibrocartilage. Although a stem-cell based approach towards regenerating the articular cartilage is attractive, cartilage generated from human mesenchymal stem cells (hMSCs) often lack the function, organization and stability of the native cartilage. Thus, there is a need to develop effective methods to engineer physiologic cartilage tissues from hMSCs in vitro and assess their outcomes in vivo. This dissertation focused on three coordinated aims: establish a simple in vivo model for studying the maturation of osteochondral tissues by showing that subcutaneous implantation in a mouse recapitulates native endochondral ossification (Aim 1), (ii) develop a robust method for engineering physiologic cartilage discs from self-assembling hMSCs (Aim 2), and (iii) improve the organization and stability of cartilage discs by implementing spatiotemporal control during induction in vitro (Aim 3). First, the usefulness of subcutaneous implantation in mice for studying the development and maintenance of osteochondral tissues in vivo was determined. By studying juvenile bovine osteochondral tissues, similarities in the profiles of endochondral ossification between the native and ectopic processes were observed. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

Inability of Low Oxygen Tension to Induce Chondrogenesis in Human Infrapatellar Fat Pad Mesenchymal Stem Cells
fcell-09-703038 July 20, 2021 Time: 15:26 # 1 ORIGINAL RESEARCH published: 26 July 2021 doi: 10.3389/fcell.2021.703038 Inability of Low Oxygen Tension to Induce Chondrogenesis in Human Infrapatellar Fat Pad Mesenchymal Stem Cells Samia Rahman1, Alexander R. A. Szojka1, Yan Liang1, Melanie Kunze1, Victoria Goncalves1, Aillette Mulet-Sierra1, Nadr M. Jomha1 and Adetola B. Adesida1,2* 1 Laboratory of Stem Cell Biology and Orthopedic Tissue Engineering, Division of Orthopedic Surgery and Surgical Research, Department of Surgery, University of Alberta, Edmonton, AB, Canada, 2 Division of Otolaryngology-Head and Neck Surgery, Department of Surgery, University of Alberta Hospital, Edmonton, AB, Canada Objective: Articular cartilage of the knee joint is avascular, exists under a low oxygen tension microenvironment, and does not self-heal when injured. Human infrapatellar fat pad-sourced mesenchymal stem cells (IFP-MSC) are an arthroscopically accessible source of mesenchymal stem cells (MSC) for the repair of articular cartilage defects. Human IFP-MSC exists physiologically under a low oxygen tension (i.e., 1–5%) Edited by: microenvironment. Human bone marrow mesenchymal stem cells (BM-MSC) exist Yi Zhang, physiologically within a similar range of oxygen tension. A low oxygen tension of Central South University, China 2% spontaneously induced chondrogenesis in micromass pellets of human BM-MSC. Reviewed by: Dimitrios Kouroupis, However, this is yet to be demonstrated in human IFP-MSC or other adipose tissue- University of Miami, United States sourced MSC. In this study, we explored the potential of low oxygen tension at 2% to Dhirendra Katti, Indian Institute of Technology Kanpur, drive the in vitro chondrogenesis of IFP-MSC. -

Bones of the Skeletal System
BIOLOGY 211: HUMAN ANATOMY & PHYSIOLOGY ********************************************************************************************************* BONES OF THE SKELETAL SYSTEM ********************************************************************************************************** Reference: Saladin, KS: Anatomy & Physiology, The Unity of Form and Function, 6th ed. (2012) or 7th ed. (2015) Please review Chapters 7 & 8 before beginning this lab. INTRODUCTION The skeletal system has a number of important functions in the human body. It is the framework around which the body is organized, it provides levers for muscles to pull against, and it surrounds and protects many soft organs. Equally important, bones serve as a "buffer" in which calcium and other ions can be deposited and withdrawn according to the changing needs of the body, and they are the site of almost all blood cell production. Contrary to our popular conceptions, bones are not rigid, inflexible structures: they are constantly changing, and can have a remarkable degree of flexibility before they break. The organs of the skeletal system are the bones and joints, and like all organs are composed of different types of tissue. Although we tend to classify them into "types" such as "long bones", "flat bones", etc., each is in fact unique and ideally suited to its particular location and function. We classify bones as belonging to either: a) the axial skeleton (head and trunk) b) the appendicular skeleton (arms and legs), However, you should always bear in mind that the entire skeletal system functions as a unit. If you look at any bone, you will see that it is rarely flat or smooth. Bones have a variety of bumps, grooves, holes, etc. which allow them to serve their specific functions.