High-Resolution Vegetation History of West Africa During the Last 145 Ka
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biodiversity and Ecology of Afromontane Rainforests with Wild Coffea Arabica L
Feyera Senbeta Wakjira (Autor) Biodiversity and ecology of Afromontane rainforests with wild Coffea arabica L. populations in Ethiopia https://cuvillier.de/de/shop/publications/2260 Copyright: Cuvillier Verlag, Inhaberin Annette Jentzsch-Cuvillier, Nonnenstieg 8, 37075 Göttingen, Germany Telefon: +49 (0)551 54724-0, E-Mail: [email protected], Website: https://cuvillier.de General introduction 1 GENERAL INTRODUCTION 1.1 Background The term biodiversity is used to convey the total number, variety and variability of living organisms and the ecological complexes in which they occur (Wilson 1988; CBD 1992; Rosenzweig 1995). The concept of biological diversity can be applied to a wide range of spatial and organization scales, including genetics, species, community, and landscape scales (Noss 1990; Austin et al. 1996; Tuomisto et al. 2003). It is becoming increasingly apparent that knowledge of the role of patterns and processes that determine diversity at different scales is at the very heart of an understanding of variation in biodiversity. Processes influencing diversity operate at different spatial and temporal scales (Rosenzweig 1995; Gaston 2000). A variety of environmental events and processes, including past evolutionary development, biogeographic processes, extinctions, and current influences govern the biodiversity of a particular site (Brown and Lomolino 1998; Gaston 2000; Ricklefs and Miller 2000). Biodiversity is valued and has been studied largely because it is used, and could be used better, to sustain and improve human well-being (WWF 1993; WCMC 1994). However, there has been a rapid decline in the biodiversity of the world during the past two to three decades (Wilson 1988; Whitmore and Sayer 1992; Lugo et al. -

The State of Biodiversity in Africa a Mid-Term Review of Progress Towards the Aichi Biodiversity Targets
THE STATE OF BIODIVERSITY IN AFRICA A MID-TERM REVIEW OF PROGRESS TOWARDS THE AICHI BIODIVERSITY TARGETS 1 3 © Neil Burgess © Neil Preparation Reproduction This study was commissioned by the Division of Environmental This publication may be reproduced for educational or Law and Conventions (DELC) of the United Nations non-profit purposes without special permission, provided Environmental Programme (UNEP) under the leadership acknowledgement to the source is made. Reuse of any figures of Ms. Elizabeth Maruma Mrema, DELC Director, and the is subject to permission from the original rights holders. No direct supervision of Ms. Kamar Yousuf, Regional Biodiversity use of this publication may be made for resale or any other Multilateral Environmental Agreement (MEA) Focal Point commercial purpose without permission in writing from UNEP. for Africa. Additional funding has been provided by the Applications for permission, with a statement of purpose and UNEP World Conservation Monitoring Centre (UNEP- extent of reproduction, should be sent to the UNEP-DELC WCMC) and the Secretariat of the Convention on Biological Director, United Nations Environment Programme, P.O. Box Diversity (SCBD). The design, printing and distribution of 30552, Nairobi 00100, Kenya. this report was enabled through the financial contribution of the European Union. Disclaimer The contents of this report do not necessarily reflect the views Citation or policies of UNEP, contributory organizations or editors. The UNEP-WCMC (2016) The State of Biodiversity in Africa: designations employed and the presentations of material in this A mid-term review of progress towards the Aichi Biodiversity report do not imply the expression of any opinion whatsoever Targets. -

Climate Forcing of Tree Growth in Dry Afromontane Forest Fragments of Northern Ethiopia: Evidence from Multi-Species Responses Zenebe Girmay Siyum1,2* , J
Siyum et al. Forest Ecosystems (2019) 6:15 https://doi.org/10.1186/s40663-019-0178-y RESEARCH Open Access Climate forcing of tree growth in dry Afromontane forest fragments of Northern Ethiopia: evidence from multi-species responses Zenebe Girmay Siyum1,2* , J. O. Ayoade3, M. A. Onilude4 and Motuma Tolera Feyissa2 Abstract Background: Climate-induced challenge remains a growing concern in the dry tropics, threatening carbon sink potential of tropical dry forests. Hence, understanding their responses to the changing climate is of high priority to facilitate sustainable management of the remnant dry forests. In this study, we examined the long-term climate- growth relations of main tree species in the remnant dry Afromontane forests in northern Ethiopia. The aim of this study was to assess the dendrochronological potential of selected dry Afromontane tree species and to study the influence of climatic variables (temperature and rainfall) on radial growth. It was hypothesized that there are potential tree species with discernible annual growth rings owing to the uni-modality of rainfall in the region. Ring width measurements were based on increment core samples and stem discs collected from a total of 106 trees belonging to three tree species (Juniperus procera, Olea europaea subsp. cuspidate and Podocarpus falcatus). The collected samples were prepared, crossdated, and analyzed using standard dendrochronological methods. The formation of annual growth rings of the study species was verified based on successful crossdatability and by correlating tree-ring widths with rainfall. Results: The results showed that all the sampled tree species form distinct growth boundaries though differences in the distinctiveness were observed among the species. -

Lesotho Fourth National Report on Implementation of Convention on Biological Diversity
Lesotho Fourth National Report On Implementation of Convention on Biological Diversity December 2009 LIST OF ABBREVIATIONS AND ACRONYMS ADB African Development Bank CBD Convention on Biological Diversity CCF Community Conservation Forum CITES Convention on International Trade in Endangered Species CMBSL Conserving Mountain Biodiversity in Southern Lesotho COP Conference of Parties CPA Cattle Post Areas DANCED Danish Cooperation for Environment and Development DDT Di-nitro Di-phenyl Trichloroethane EA Environmental Assessment EIA Environmental Impact Assessment EMP Environmental Management Plan ERMA Environmental Resources Management Area EMPR Environmental Management for Poverty Reduction EPAP Environmental Policy and Action Plan EU Environmental Unit (s) GA Grazing Associations GCM Global Circulation Model GEF Global Environment Facility GMO Genetically Modified Organism (s) HIV/AIDS Human Immuno Virus/Acquired Immuno-Deficiency Syndrome HNRRIEP Highlands Natural Resources and Rural Income Enhancement Project IGP Income Generation Project (s) IUCN International Union for Conservation of Nature and Natural Resources LHDA Lesotho Highlands Development Authority LMO Living Modified Organism (s) Masl Meters above sea level MDTP Maloti-Drakensberg Transfrontier Conservation and Development Project MEAs Multi-lateral Environmental Agreements MOU Memorandum Of Understanding MRA Managed Resource Area NAP National Action Plan NBF National Biosafety Framework NBSAP National Biodiversity Strategy and Action Plan NEAP National Environmental Action -

Comparative Wood Anatomy of Afromontane and Bushveld Species from Swaziland, Southern Africa
IAWA Bulletin n.s., Vol. 11 (4), 1990: 319-336 COMPARATIVE WOOD ANATOMY OF AFROMONTANE AND BUSHVELD SPECIES FROM SWAZILAND, SOUTHERN AFRICA by J. A. B. Prior 1 and P. E. Gasson 2 1 Department of Biology, Imperial College of Science, Technology & Medicine, London SW7 2BB, U.K. and 2Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3DS, U.K. Summary The habit, specific gravity and wood anat of the archaeological research, uses all the omy of 43 Afromontane and 50 Bushveld well preserved, qualitative anatomical charac species from Swaziland are compared, using ters apparent in the charred modem samples qualitative features from SEM photographs in an anatomical comparison between the of charred samples. Woods with solitary ves two selected assemblages of trees and shrubs sels, scalariform perforation plates and fibres growing in areas of contrasting floristic com with distinctly bordered pits are more com position. Some of the woods are described in mon in the Afromontane species, whereas Kromhout (1975), others are of little com homocellular rays and prismatic crystals of mercial importance and have not previously calcium oxalate are more common in woods been investigated. Few ecological trends in from the Bushveld. wood anatomical features have previously Key words: Swaziland, Afromontane, Bush been published for southern Africa. veld, archaeological charcoal, SEM, eco The site of Sibebe Hill in northwest Swazi logical anatomy. land (26° 15' S, 31° 10' E) (Price Williams 1981), lies at an altitude of 1400 m, amidst a Introduction dramatic series of granite domes in the Afro Swaziland, one of the smallest African montane forest belt (White 1978). -

The Vulnerability of Indo-Pacific Mangrove Forests to Sea-Level Rise
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln USGS Staff -- Published Research US Geological Survey 10-22-2015 The vulnerability of Indo-Pacific mangrove forests to sea-level rise Catherine E. Lovelock The University of Queensland Donald R. Cahoon United States Geological Survey, [email protected] Daniel A. Friess National University of Singapore Glenn R. Guntenspergen United States Geological Survey, [email protected] Ken W. Krauss United States Geological Survey See next page for additional authors Follow this and additional works at: https://digitalcommons.unl.edu/usgsstaffpub Part of the Geology Commons, Oceanography and Atmospheric Sciences and Meteorology Commons, Other Earth Sciences Commons, and the Other Environmental Sciences Commons Lovelock, Catherine E.; Cahoon, Donald R.; Friess, Daniel A.; Guntenspergen, Glenn R.; Krauss, Ken W.; Reef, Ruth; Rogers, Kerrylee; Saunders, Megan L.; Sidik, Frida; Swales, Andrew; Saintilan, Neil; Thuyen, Le Xuan; and Triet, Tran, "The vulnerability of Indo-Pacific mangrove forests to sea-level rise" (2015). USGS Staff -- Published Research. 988. https://digitalcommons.unl.edu/usgsstaffpub/988 This Article is brought to you for free and open access by the US Geological Survey at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in USGS Staff -- Published Research by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Authors Catherine E. Lovelock, Donald R. Cahoon, Daniel A. Friess, Glenn R. Guntenspergen, Ken W. Krauss, Ruth Reef, Kerrylee Rogers, Megan L. Saunders, Frida Sidik, Andrew Swales, Neil Saintilan, Le Xuan Thuyen, and Tran Triet This article is available at DigitalCommons@University of Nebraska - Lincoln: https://digitalcommons.unl.edu/ usgsstaffpub/988 LETTER doi:10.1038/nature15538 The vulnerability of Indo-Pacific mangrove forests to sea-level rise Catherine E. -
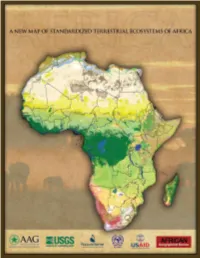
A New Map of Standardized Terrestrial Ecosystems of Africa
Major contributors to this publication include: The Association of American Geographers is a nonprofit scientific and educational society with a membership of over 10,500 individuals from more than 60 countries. AAG members are geographers and related professionals who work in the public, private, and academic sectors to advance the theory, methods, and practice of geography. This booklet is published by AAG as a special supplement to the African Geographical Review. The U.S. Geological Survey (USGS) was created in 1879 as a science agency charged with providing information and understanding to help resolve complex natural resource problems across the nation and around the world. The mission of the USGS is to provide relevant, impartial scientific information to 1) describe and understand the Earth, 2) minimize loss of life and property from natural disasters, 3) manage water, biological, energy, and mineral resources, and 4) enhance and protect our quality of life. NatureServe is an international conservation nonprofit dedicated to providing the sci- entific basis for effective conservation action. NatureServe’s network in the Americas includes more than 80 member institutions that collect and maintain a unique body of scientific knowledge about the species and ecosystems. The information products, data management tools, and biodiversity expertise that NatureServe’s scientists, technologists, and other professionals provide help meet local, national, and global conservation needs. The Regional Centre for Mapping of Resources for Development (RCMRD) was es- tablished in Nairobi, Kenya in 1975 to provide quality Geo-Information and allied Information Technology products and services in environmental and resource manage- ment for sustainable development in our member countries and beyond. -

Distribution, Habitat Structure and Troop Size in Eastern Cape
Distribution, habitat structure and troop size in Eastern Cape samango monkeys Cercopithecus albogularis labiatus (Primates: Cercopithecoidea) A dissertation submitted to the Department of Zoology and Entomology in fulfilment of the requirements for the degree of Master of Science in Zoology the University of Fort Hare by Vusumzi Martins Supervisors: Dr Fabien Génin and Prof. Judith Masters GENERAL ABSTRACT The samango monkey subspecies Cercopithecus albogularis labiatus is endemic to South Africa, and known to occur in Afromontane forests. There has been a major decline in this subspecies, exceeding 30% in some populations over the past 30 years, primarily as a result of the loss of suitable habitat. A second samango subspecies, C. a. erythrarchus, occurs near the northern border of South Africa, mainly in coastal lowland forest, and the distributions of the two subspecies do not overlap. C. a. labiatus was thought to be confined to Afromontane forests, but the study described here focused on C. a. labiatus populations that were recently identified in the Indian Ocean Belt forests near East London. I undertook to assess the distribution of C. a. labiatus in the Eastern Cape, to evaluate the habitat structures of the Afromontane and Indian Ocean coastal belt forests, and to understand the effect these habitats have on essential aspects of the socio-ecology of the C. a. labiatus populations. Distribution surveys were conducted in protected areas throughout the Eastern Cape, and samango monkeys were found to be present within forest patches in the Amatola Mountains, Eastern Cape dune forests and the Transkei coastal scarp forests. The sizes and composition of two troops were assessed: one troop in the Amatole forests and one in the Eastern Cape dune forests. -

Florida Keys National Marine Sanctuary Water Quality
FLORIDA KEYS NATIONAL MARINE SANCTUARY WATER QUALITY PROTECTION PROGRAM STEERING COMMITTEE MEETING January 25, 2012 MARATHON GOVERNMENT CENTER, 2798 Overseas Highway, Marathon, FL 33050 MINUTES Members Present: Tracy Ziegler, Everglades and Dry Tortugas National Parks Charles Brooks, Key Largo Wastewater Treatment District Armando Rilaboy, South Florida Water Management District Susan Hammaker, Key Largo Wastewater Treatment District, Sanctuary Advisory Council Ron Sutton, Key Colony Beach Pete Worthington, City of Marathon Charles Briggs, Department of Health, Bureau of Onsite Sewage Treatment Systems George Neugent, Monroe County Commissioner Charlie Causey Florida Keys Environmental Fund Sandy Walters, SWC, Inc., representing maritime interests of the Florida Keys Judy Cheon, Florida Keys Aqueduct Authority Gil McRae, Florida Fish and Wildlife Commission, Fish and Wildlife Research Institute Chris Bergh, The Nature Conservancy Bruce Popham, Sanctuary Advisory Council and a member of the business community Billy Causey, Southeast Region of National Marine Sanctuaries Anne Morkill, Fish and Wildlife Service, Florida Keys National Wildlife Refuges I. Call Meeting to Order Welcoming George Garrett George Garrett addressed the committee and welcomed everyone, including the EPA regional administrator region IV, Ms. Gwedolyn Keyes Fleming. She is always welcome to attend the meetings. Richard Harvey thanked George and introduced Ms. Gwen Keyes Fleming and the other guests, Mr. Jim Giattina, Water Protection Division Director, and Brandi Jenkins, Special Assistant to the regional administrator and Javoyne Hicks White, Chief of Staff for the regional administrator. He explained that the regional director will be making a few remarks. She has participated in a few meetings since she has been here. Jon Iglehart will make a statement on behalf of Secretary Vinyard, who could not be here today. -

Ushaka Sea World Education Mangrove Ecosystem Study
uShaka Sea World Education Grade 10 Life Sciences Mangrove Ecosystem Study 1 Mangrove Ecosystem Study In this study you will be looking at the mangroves as an ecosystem. Your study will deal with all the following: • abiotic and biotic factors and the interactions between them. • trophic relationships in that ecosystem. • the changes between high and low tide. • biodiversity within the ecosystem using information keys and food webs. • positive and/or negative human impact/influence on the ecosystem. Curriculum link GRADE 10 STRAND 3: ENVIRONMENTAL STUDIES - Biosphere to Ecosystems Aquatic biomes of Southern Africa: How climate, soils and vegetation influence the organisms found in each biome. Ecosystem structure and functioning and the abiotic and biotic factors and how they affect the community: Abiotic factors: •physiographic factors (aspect, slope, altitude) • light (day length, seasonal changes) • temperature (effect of day/night, seasons) • water (water cycle, importance of wetlands) Biotic factors: (Links to Grade 8) • producers • consumers • decomposers Energy flow through ecosystems and relationship to trophic structure (food pyramids): Producers, consumers (herbivores, carnivores and omnivores, decomposers). Human activities in and interactions with the natural environment. 2 READING South African Mangrove Forests Together with coral reefs and seagrass beds, mangroves rank as one of the most productive ecosystems in the world. Physical Environment Mangrove forests are found in the littoral zone, the band between the sea and the land that is flooded and exposed during the rise and fall of the tide. They occur in estuaries which are either predominantly or permanently open to the sea and tidal in nature. Mangroves are usually only found in tropical climates, as they need consistent warm conditions for development and survival. -

The Indo Pacific Forest C Study Handout
The Indo-Pacific Forest Carbon Study A collaborative study with the US Forest Service, Institute of Pacific Islands Forestry, International Programs, and the Center for International Forest Research Recent work by IPIF scientists have found that the C pools of mangroves exceed that of upland tropical forests because they exist on C-rich peat soils and have relative high levels of aboveground biomass (Fig. 1). Mangrove forests are widely distributed in the western Pacific and have been subject to high rates of land cover change related to aquaculture and Yap Crew coastal development. Southeast Indo-Pacific Forest Carbon Study Asia supports the world’s largest October 25, 2008 areas of mangrove originally extending over 6.8 million ha representing 34-42% if the world’s total. These forests are vulnerable to land cover change and global change especially sea level rise. Because of the many ecosystem services they provide and the high quantity of C they store, they Ecosystem C pools of Selected Tropical Forests would be ideal candidates for REDD 800 strategies. 700 600 500 Scientists at the center for International Aboveground 400 Forestry Research (CIFOR) have reported Belowground 300 that tropical forests over peat soils in the 200 C Mass (Mg/ha) Mass C Asian Pacific also contain immense C pools 100 that when subjected to land cover change 0 TDF Mex TMF TWF Mont rain Mangrove Mangrove are significant sources of anthropogenic Brasil Mex CR Palau Yap emissions of CO 2 and CH 4. Yet little Soils only to a 1m depth- Palau mangrove mass to 160 cm was ~963 quantitative work exists in accurately Mg/ha and at Yap mass to 231 cm was 1,232Mg/ha.From Jaramillo ascertaining the C pools of undisturbed et al. -

Biodiversity in Sub-Saharan Africa and Its Islands Conservation, Management and Sustainable Use
Biodiversity in Sub-Saharan Africa and its Islands Conservation, Management and Sustainable Use Occasional Papers of the IUCN Species Survival Commission No. 6 IUCN - The World Conservation Union IUCN Species Survival Commission Role of the SSC The Species Survival Commission (SSC) is IUCN's primary source of the 4. To provide advice, information, and expertise to the Secretariat of the scientific and technical information required for the maintenance of biologi- Convention on International Trade in Endangered Species of Wild Fauna cal diversity through the conservation of endangered and vulnerable species and Flora (CITES) and other international agreements affecting conser- of fauna and flora, whilst recommending and promoting measures for their vation of species or biological diversity. conservation, and for the management of other species of conservation con- cern. Its objective is to mobilize action to prevent the extinction of species, 5. To carry out specific tasks on behalf of the Union, including: sub-species and discrete populations of fauna and flora, thereby not only maintaining biological diversity but improving the status of endangered and • coordination of a programme of activities for the conservation of bio- vulnerable species. logical diversity within the framework of the IUCN Conservation Programme. Objectives of the SSC • promotion of the maintenance of biological diversity by monitoring 1. To participate in the further development, promotion and implementation the status of species and populations of conservation concern. of the World Conservation Strategy; to advise on the development of IUCN's Conservation Programme; to support the implementation of the • development and review of conservation action plans and priorities Programme' and to assist in the development, screening, and monitoring for species and their populations.