Phylogeny of Lampridiform Fishes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CHECKLIST and BIOGEOGRAPHY of FISHES from GUADALUPE ISLAND, WESTERN MEXICO Héctor Reyes-Bonilla, Arturo Ayala-Bocos, Luis E
ReyeS-BONIllA eT Al: CheCklIST AND BIOgeOgRAphy Of fISheS fROm gUADAlUpe ISlAND CalCOfI Rep., Vol. 51, 2010 CHECKLIST AND BIOGEOGRAPHY OF FISHES FROM GUADALUPE ISLAND, WESTERN MEXICO Héctor REyES-BONILLA, Arturo AyALA-BOCOS, LUIS E. Calderon-AGUILERA SAúL GONzáLEz-Romero, ISRAEL SáNCHEz-ALCántara Centro de Investigación Científica y de Educación Superior de Ensenada AND MARIANA Walther MENDOzA Carretera Tijuana - Ensenada # 3918, zona Playitas, C.P. 22860 Universidad Autónoma de Baja California Sur Ensenada, B.C., México Departamento de Biología Marina Tel: +52 646 1750500, ext. 25257; Fax: +52 646 Apartado postal 19-B, CP 23080 [email protected] La Paz, B.C.S., México. Tel: (612) 123-8800, ext. 4160; Fax: (612) 123-8819 NADIA C. Olivares-BAñUELOS [email protected] Reserva de la Biosfera Isla Guadalupe Comisión Nacional de áreas Naturales Protegidas yULIANA R. BEDOLLA-GUzMáN AND Avenida del Puerto 375, local 30 Arturo RAMíREz-VALDEz Fraccionamiento Playas de Ensenada, C.P. 22880 Universidad Autónoma de Baja California Ensenada, B.C., México Facultad de Ciencias Marinas, Instituto de Investigaciones Oceanológicas Universidad Autónoma de Baja California, Carr. Tijuana-Ensenada km. 107, Apartado postal 453, C.P. 22890 Ensenada, B.C., México ABSTRACT recognized the biological and ecological significance of Guadalupe Island, off Baja California, México, is Guadalupe Island, and declared it a Biosphere Reserve an important fishing area which also harbors high (SEMARNAT 2005). marine biodiversity. Based on field data, literature Guadalupe Island is isolated, far away from the main- reviews, and scientific collection records, we pres- land and has limited logistic facilities to conduct scien- ent a comprehensive checklist of the local fish fauna, tific studies. -

Early Stages of Fishes in the Western North Atlantic Ocean Volume
ISBN 0-9689167-4-x Early Stages of Fishes in the Western North Atlantic Ocean (Davis Strait, Southern Greenland and Flemish Cap to Cape Hatteras) Volume One Acipenseriformes through Syngnathiformes Michael P. Fahay ii Early Stages of Fishes in the Western North Atlantic Ocean iii Dedication This monograph is dedicated to those highly skilled larval fish illustrators whose talents and efforts have greatly facilitated the study of fish ontogeny. The works of many of those fine illustrators grace these pages. iv Early Stages of Fishes in the Western North Atlantic Ocean v Preface The contents of this monograph are a revision and update of an earlier atlas describing the eggs and larvae of western Atlantic marine fishes occurring between the Scotian Shelf and Cape Hatteras, North Carolina (Fahay, 1983). The three-fold increase in the total num- ber of species covered in the current compilation is the result of both a larger study area and a recent increase in published ontogenetic studies of fishes by many authors and students of the morphology of early stages of marine fishes. It is a tribute to the efforts of those authors that the ontogeny of greater than 70% of species known from the western North Atlantic Ocean is now well described. Michael Fahay 241 Sabino Road West Bath, Maine 04530 U.S.A. vi Acknowledgements I greatly appreciate the help provided by a number of very knowledgeable friends and colleagues dur- ing the preparation of this monograph. Jon Hare undertook a painstakingly critical review of the entire monograph, corrected omissions, inconsistencies, and errors of fact, and made suggestions which markedly improved its organization and presentation. -

A New Species of Cretaceous Acanthomorph from Canada 15 February 2016, by Sarah Gibson
A new species of Cretaceous acanthomorph from Canada 15 February 2016, by Sarah Gibson to sit flush along the body, helping the fish swim faster by reducing drag, or they can be extended completely out to act as a defense mechanism, in case you are a predator looking for a quick bite. Near the base of the Acanthomorpha phylogenetic tree is a small group of fishes, Polymixiiformes, comprised of a single living genus, Polymixia, more commonly known as the beardfish. This innocuous fish seems harmless, but according to many ichthyologists, Polymixia is just one key to understanding acanthomorph relationships. Unraveling the evolutionary relationships is difficult with a single living genus, but thankfully, polymixiiforms have a fossil record dating back to the Cretaceous, containing an increasing number of taxa as new discoveries are being made, particularly in deposits in North America, where fewer acanthomorph fossils are known compared to Figuring out fish relationships is no small feat. Credit: the more-studied Eastern Tethys Ocean deposits in Near et al. 2013 Europe. For being one of the largest groups of vertebrates, and having one of the richer fossil records among organisms, the relationships of fishes are still hotly debated. Humongous datasets are being compiled that involve molecular (both nuclear and mitochondrial) data, compared and contrasted with thorough morphological analyses. (I'm not going to get into all of it here, simply because of its sheer complexity.) What I am going to get into, however, is the fossil record of one subset of fishes, the acanthomorphs. The stout beardfish, Polymixia nobilis. Credit: Wikipedia Acanthomorphs are teleost fishes that possess true fin spines: a set of prominent, sharp, unsegmented spines in the front portion of their dorsal and/or anal fins, followed by a portion of One such new species was recently described by pliable, segmented, "softer" looking rays. -

Crestfish Lophotus Lacepede (Giorna, 1809) and Scalloped Ribbonfish Zu Cristatus (Bonelli, 1819) in the Northern Coast of Sicily, Italy
ISSN: 0001-5113 ACTA ADRIAT., ORIGINAL SCIENTIFIC PAPER AADRAY 58(1): 137 - 146, 2017 Occurrence of two rare species from order Lampriformes: Crestfish Lophotus lacepede (Giorna, 1809) and scalloped ribbonfish Zu cristatus (Bonelli, 1819) in the northern coast of Sicily, Italy Fabio FALSONE1, Michele Luca GERACI1, Danilo SCANNELLA1, Charles Odilichukwu R. OKPALA1, Giovan Battista GIUSTO1, Mar BOSCH-BELMAR2, Salvatore GANCITANO1 and Gioacchino BONO1 1Institute for the Coastal Marine Environment, IAMC‑CNR, 91026 Mazara del Vallo, Sicily, Italy 2Consorzio Nazionale Interuniversitario per le Scienze del Mare (CoNISMa), Rome, Italy Corresponding author, e‑mail: [email protected] The bony fish Lophotus lacepede (Giorna, 1809) and Zu cristatus (Bonelli, 1819) are the two species rarely recorded within the Mediterranean basin, usually reported as accidentally captured in depth (mesopelagic) fishing operations. In the current work, we present the first record of L. lacepede and Z. cristatus in fishing catches from southwestern Tyrrhenian Sea. Moreover, in order to improve existent biological/ecological knowledge, some bio-related aspects such as feeding aspect, sexual maturity and age estimate have been discussed. Key words: crestfish, scalloped ribbonfish, meristic features, vertebrae, growth ring INTRODUCTION species of Lophotidae family, the L. lacepede inhabits the epipelagic zone, although it could The target species of this study (Lophotus also be recorded in most oceans from the surface lacepede and Zu cristatus) belong to Lophotidae up to 1000 m depth (HEEMSTRA, 1986; PALMER, (Bonaparte, 1845) and Trachipteridae (Swain- 1986; OLNEY, 1999). First record of this spe- son, 1839) families respectively, including the cies in the Mediterranean Basin was from the Lampriformes order (consisted of 7 families). -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

Percomorph Phylogeny: a Survey of Acanthomorphs and a New Proposal
BULLETIN OF MARINE SCIENCE, 52(1): 554-626, 1993 PERCOMORPH PHYLOGENY: A SURVEY OF ACANTHOMORPHS AND A NEW PROPOSAL G. David Johnson and Colin Patterson ABSTRACT The interrelationships of acanthomorph fishes are reviewed. We recognize seven mono- phyletic terminal taxa among acanthomorphs: Lampridiformes, Polymixiiformes, Paracan- thopterygii, Stephanoberyciformes, Beryciformes, Zeiformes, and a new taxon named Smeg- mamorpha. The Percomorpha, as currently constituted, are polyphyletic, and the Perciformes are probably paraphyletic. The smegmamorphs comprise five subgroups: Synbranchiformes (Synbranchoidei and Mastacembeloidei), Mugilomorpha (Mugiloidei), Elassomatidae (Elas- soma), Gasterosteiformes, and Atherinomorpha. Monophyly of Lampridiformes is justified elsewhere; we have found no new characters to substantiate the monophyly of Polymixi- iformes (which is not in doubt) or Paracanthopterygii. Stephanoberyciformes uniquely share a modification of the extrascapular, and Beryciformes a modification of the anterior part of the supraorbital and infraorbital sensory canals, here named Jakubowski's organ. Our Zei- formes excludes the Caproidae, and characters are proposed to justify the monophyly of the group in that restricted sense. The Smegmamorpha are thought to be monophyletic principally because of the configuration of the first vertebra and its intermuscular bone. Within the Smegmamorpha, the Atherinomorpha and Mugilomorpha are shown to be monophyletic elsewhere. Our Gasterosteiformes includes the syngnathoids and the Pegasiformes -

First Record of Polka-Dot Ribbonfish from India 3
Marine Biodiversity Records, page 1 of 4. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S1755267211001151; Vol. 5; e8; 2012 Published online First record of Polka-dot ribbonfish Desmodema polystictum (Pisces: Trachipteridae) from Indian waters p.u. zacharia and k. kannan Central Marine Fisheries Research Institute, PB No.1603, Ernakulam North PO, Cochin-682 018, Kerala, India Polka-dot ribbonfish Desmodema polystictum was recorded for the first time from Indian waters. A single specimen of D. polystictum (107 cm total length and weighing 480 g) was collected from Tharuvaikulam landing centre, north to Tuticorin, on the south-east coast of India during September 2010. The distinguishing characters of the species from other species of the family are discussed. Morphometric and meristic characters of D. polystictum are presented in this paper. With the present report, the distribution area of this species now extends to the Indian waters. Keywords: first record, Polka-dot ribbonfish, Desmodema polystictum, Indian waters Submitted 6 September 2011; accepted 29 November 2011 INTRODUCTION of D. polystictum aresilverywithprofusedarkspotting(polka dotted) but the adults lack spots. The fish of the family Trachipteridae are characterized by long Froese & Pauly (2010) state that Desmodema polystictum compressed ribbon or tape-shaped body, short head, and probably has a circumtropical distribution; it was reported narrow mouth (Heemstra & Kannemeyer, 1986). The pectoral from Japan, Taiwan (Shen, 1993), Philippines, Australia, fin is small, pelvic long and fan like in young composed of New Zealand (Paulin et al., 1989); Western Pacific (Ogilby, several rays, absent in adults. Anal fin absent. These fish also 1897) and 16811′N to Namibia (Aguiar & Que´ro, 1990); have a high dorsal fin that actually occupies the entire length South Africa in the eastern Atlantic; Florida, USA (Moore of its back with origin well behind tip of snout. -

The Phylogenetic Relationships of Lampridiform Fishes (Teleostei: Acanthomorpha), Based on a Total-Evidence Analysis of Morphological and Molecular Data E
MOLECULAR PHYLOGENETICS AND EVOLUTION Vol. 10, No. 3, December, pp. 417–425, 1998 ARTICLE NO. FY980532 The Phylogenetic Relationships of Lampridiform Fishes (Teleostei: Acanthomorpha), Based on a Total-Evidence Analysis of Morphological and Molecular Data E. O. Wiley,*,† G. David Johnson,† and Walter Wheaton Dimmick* *Natural History Museum and Department of Systematics and Ecology, University of Kansas, Lawrence, Kansas 66045; and †Division of Fishes, National Museum of Natural History, Washington, DC 20560 Received November 11, 1997; revised March 10, 1998 The Lampridiformes comprise 21 species in 12 gen- We investigated the phylogenetic relationships era and 7 families. The relatively small (40 cm) velifer- among five species of lampridiform fishes, three basal ids and much larger (1.8 m) opahs (Lamprididae) are outgroup species (two aulopiforms and one myctophi- deep bodied. The remaining families are elongate and form), and two species of non-lampridiform acantho- range in body size from the small tube-eyes (Stylephori- morphs (Polymixia and Percopsis) using a combined dae, 31 cm) to the very large Regalecus (17 m). All but parsimony analysis of morphological and molecular the near shore veliferids are oceanic fishes, ranging data. Morphological characters included 28 transfor- mation series obtained from the literature. Molecular from the epipelagic realm to abyssal depths. Lampridi- characters included 223 informative transformation form fishes are characterized by a uniquely protusible series from an aligned 854-base pair fragment of 12S upper jaw apparatus and have been considered a mtDNA and 139 informative transformation series from natural group since Regan (1907) named them. Prior to an aligned 561-base pair fragment of 16S mtDNA. -

Euteleostei: Aulopiformes) and the Timing of Deep-Sea Adaptations ⇑ Matthew P
Molecular Phylogenetics and Evolution 57 (2010) 1194–1208 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Estimating divergence times of lizardfishes and their allies (Euteleostei: Aulopiformes) and the timing of deep-sea adaptations ⇑ Matthew P. Davis a, , Christopher Fielitz b a Museum of Natural Science, Louisiana State University, 119 Foster Hall, Baton Rouge, LA 70803, USA b Department of Biology, Emory & Henry College, Emory, VA 24327, USA article info abstract Article history: The divergence times of lizardfishes (Euteleostei: Aulopiformes) are estimated utilizing a Bayesian Received 18 May 2010 approach in combination with knowledge of the fossil record of teleosts and a taxonomic review of fossil Revised 1 September 2010 aulopiform taxa. These results are integrated with a study of character evolution regarding deep-sea evo- Accepted 7 September 2010 lutionary adaptations in the clade, including simultaneous hermaphroditism and tubular eyes. Diver- Available online 18 September 2010 gence time estimations recover that the stem species of the lizardfishes arose during the Early Cretaceous/Late Jurassic in a marine environment with separate sexes, and laterally directed, round eyes. Keywords: Tubular eyes have arisen independently at different times in three deep-sea pelagic predatory aulopiform Phylogenetics lineages. Simultaneous hermaphroditism evolved a single time in the stem species of the suborder Character evolution Deep-sea Alepisauroidei, the clade of deep-sea aulopiforms during the Early Cretaceous. This result indicates the Euteleostei oldest known evolutionary event of simultaneous hermaphroditism in vertebrates, with the Alepisauroidei Hermaphroditism being the largest vertebrate clade with this reproductive strategy. Divergence times Ó 2010 Elsevier Inc. -

First Record of Polka-Dot Ribbonfish Desmodema Polystictum (Pisces
Marine Biodiversity Records, page 1 of 4. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S1755267211001151; Vol. 5; e8; 2012 Published online First record of Polka-dot ribbonfish Desmodema polystictum (Pisces: Trachipteridae) from Indian waters p.u. zacharia and k. kannan Central Marine Fisheries Research Institute, PB No.1603, Ernakulam North PO, Cochin-682 018, Kerala, India Polka-dot ribbonfish Desmodema polystictum was recorded for the first time from Indian waters. A single specimen of D. polystictum (107 cm total length and weighing 480 g) was collected from Tharuvaikulam landing centre, north to Tuticorin, on the south-east coast of India during September 2010. The distinguishing characters of the species from other species of the family are discussed. Morphometric and meristic characters of D. polystictum are presented in this paper. With the present report, the distribution area of this species now extends to the Indian waters. Keywords: first record, Polka-dot ribbonfish, Desmodema polystictum, Indian waters Submitted 6 September 2011; accepted 29 November 2011 INTRODUCTION of D. polystictum aresilverywithprofusedarkspotting(polka dotted) but the adults lack spots. The fish of the family Trachipteridae are characterized by long Froese & Pauly (2010) state that Desmodema polystictum compressed ribbon or tape-shaped body, short head, and probably has a circumtropical distribution; it was reported narrow mouth (Heemstra & Kannemeyer, 1986). The pectoral from Japan, Taiwan (Shen, 1993), Philippines, Australia, fin is small, pelvic long and fan like in young composed of New Zealand (Paulin et al., 1989); Western Pacific (Ogilby, several rays, absent in adults. Anal fin absent. These fish also 1897) and 16811′N to Namibia (Aguiar & Que´ro, 1990); have a high dorsal fin that actually occupies the entire length South Africa in the eastern Atlantic; Florida, USA (Moore of its back with origin well behind tip of snout. -
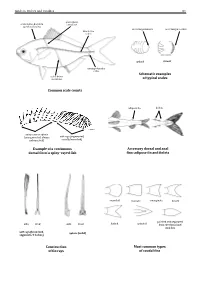
Field Identification Guide to the Living Marine Resources in Kenya
Guide to Orders and Families 81 lateral line scales above scales before dorsal fin outer margin smooth outer margin toothed (predorsal scales) lateral–line 114 scales cycloid ctenoidِّ scales circumpeduncular Schematic examples lateral line of typical scales scales below Common scale counts adipose fin finlets soft rays (segmented, spinyunbranched) rays or spines usually branched) (unsegmented, always Example of a continuous Accessory dorsal and anal dorsal fin of a spiny–rayed fish fins: adipose fin and finlets rounded truncate emarginate lunate side front side front from the dorsal and pointed and separated forked pointed soft rays (branched, spines (solid) segments, 2 halves) anal fins Construction Most common types of fin rays of caudal fins 82 Bony Fishes GUIDE TO ORDERS AND FAMILIES Order ELOPIFORMES – Tarpons and allies Fin spines absent; a single dorsal fin located above middle of body; pelvic fins in abdominal position; lateral line present; 23–25 branchiostegal rays; upper jaw extending past eye; tip of snout not overhanging mouth; colour silvery. ELOPIDAE Page 121 very small scales Ladyfishes To 90 cm. Coastal marine waters and estuaries; pelagic. A single species included in the Guide to Species.underside of head large mouth gular plate MEGALOPIDAE Page 121 last ray long Tarpons large scales To 55 cm. Coastal marine waters and estuaries; pelagic. A single species included in the Guide to Species.underside of head gular plate Order ALBULIFORMES – Bonefishes Fin spines absent; a single dorsal fin located above middle of body; pelvic fins in abdominal position; lateral line present; 6–16 branchiostegal rays; upper jaw not extending as far as front of eye; tip of snout overhanging mouth; colour silvery. -

Guide to the Coastal Marine Fishes of California
STATE OF CALIFORNIA THE RESOURCES AGENCY DEPARTMENT OF FISH AND GAME FISH BULLETIN 157 GUIDE TO THE COASTAL MARINE FISHES OF CALIFORNIA by DANIEL J. MILLER and ROBERT N. LEA Marine Resources Region 1972 ABSTRACT This is a comprehensive identification guide encompassing all shallow marine fishes within California waters. Geographic range limits, maximum size, depth range, a brief color description, and some meristic counts including, if available: fin ray counts, lateral line pores, lateral line scales, gill rakers, and vertebrae are given. Body proportions and shapes are used in the keys and a state- ment concerning the rarity or commonness in California is given for each species. In all, 554 species are described. Three of these have not been re- corded or confirmed as occurring in California waters but are included since they are apt to appear. The remainder have been recorded as occurring in an area between the Mexican and Oregon borders and offshore to at least 50 miles. Five of California species as yet have not been named or described, and ichthyologists studying these new forms have given information on identification to enable inclusion here. A dichotomous key to 144 families includes an outline figure of a repre- sentative for all but two families. Keys are presented for all larger families, and diagnostic features are pointed out on most of the figures. Illustrations are presented for all but eight species. Of the 554 species, 439 are found primarily in depths less than 400 ft., 48 are meso- or bathypelagic species, and 67 are deepwater bottom dwelling forms rarely taken in less than 400 ft.