Innervation of the Heart and Ductus Arteriosus and Other Features of the Autonomic Nervous System of the Full Term Human Fetus
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Variations of Thoracic Splanchnic Nerves and Its Clinical Implications
Int. J. Morphol., 23(3):247-251, 2005. Variations of Thoracic Splanchnic Nerves and its Clinical Implications Variaciones de los Nervios Esplácnicos Torácicos y sus Implicancias Clínicas *Tony George Jacob; ** Surbhi Wadhwa; ***Shipra Paul & ****Srijit Das JACOB, G. T.; WADHWA, S.; PAUL, S. & DAS, S. Variations of thoracic splanchnic nerves and its clinical implications. Int. J. Morphol., 23(3):247-251, 2005. SUMMARY:The present study reports an anomalous branching pattern of the thoracic sympathetic chain. At the level of T3 ganglion, an anomalous branch i.e accessory sympathetic chain (ASC) descended anteromedial to the main sympathetic chain (MSC). The MSC and the ASC communicated with each other at the level of T9, T10 and T11 ganglion, indicating the absence of classical pattern of greater, lesser and least splanchnic nerves on the right side. However, on the left side, the sympathetic chain displayed normal branching pattern. We opine that the ASC may be representing a higher origin of greater splanchnic nerve at the level of T3 ganglion and the branches from MSC at T9, T10 and T11 ganglion may be the lesser and least splanchnic nerves, which further joined the ASC (i.e presumably the greater splanchnic nerve) to form a common trunk. This common trunk pierced the right crus of diaphragm to reach the right suprarenal plexus after giving few branches to the celiac plexus. Awareness and knowledge of such anatomical variants of thoracic sympathetic chain may be helpful to surgeons in avoiding any incomplete denervation or preventing any inadvertent injury during thoracic sympathectomy. KEY WORDS: Splanchnic nerves; Sympathetic chain; Trunk thoracic; Ganglion. -

Of the Pediatric Mediastinum
MRI of the Pediatric Mediastinum Dianna M. E. Bardo, MD Director of Body MR & Co-Director of the 3D Innovation Lab Disclosures Consultant & Speakers Bureau – honoraria Koninklijke Philips Healthcare N V Author – royalties Thieme Publishing Springer Publishing Mediastinum - Anatomy Superior Mediastinum thoracic inlet to thoracic plane thoracic plane to diaphragm Inferior Mediastinum lateral – pleural surface anterior – sternum posterior – vertebral bodies Mediastinum - Anatomy Anterior T4 Mediastinum pericardium to sternum Middle Mediastinum pericardial sac Posterior Mediastinum vertebral bodies to pericardium lateral – pleural surface superior – thoracic inlet inferior - diaphragm Mediastinum – MR Challenges Motion Cardiac ECG – gating/triggering Breathing Respiratory navigation Artifacts Intubation – LMA Surgical / Interventional materials Mediastinum – MR Sequences ECG gated/triggered sequences SSFP – black blood SE – IR – GRE Non- ECG gated/triggered sequences mDIXON (W, F, IP, OP), eTHRIVE, turbo SE, STIR, DWI Respiratory – triggered, radially acquired T2W MultiVane, BLADE, PROPELLER Mediastinum – MR Sequences MRA / MRV REACT – non Gd enhanced Gd enhanced sequences THRIVE, mDIXON, mDIXON XD Mediastinum – Contents Superior Mediastinum PVT Left BATTLE: Phrenic nerve Vagus nerve Structures at the level of the sternal angle Thoracic duct Left recurrent laryngeal nerve (not the right) CLAPTRAP Brachiocephalic veins Cardiac plexus Aortic arch (and its 3 branches) Ligamentum arteriosum Thymus Aortic arch (inner concavity) Trachea Pulmonary -

Association of Arrhythmia in Patients with Cervical Spondylosis: a Nationwide Population-Based Cohort Study
Journal of Clinical Medicine Article Association of Arrhythmia in Patients with Cervical Spondylosis: A Nationwide Population-Based Cohort Study Shih-Yi Lin 1,2, Wu-Huei Hsu 1,3, Cheng-Chieh Lin 1,4, Cheng-Li Lin 5,6, Chun-Hao Tsai 1,7, Chih-Hsueh Lin 1,4, Der-Cherng Chen 7, Tsung-Chih Lin 8, Chung-Y. Hsu 1 and Chia-Hung Kao 1,9,10,* ID 1 Graduate Institute of Biomedical Sciences and School of Medicine, College of Medicine, China Medical University, No. 2, Yuh-Der Road, Taichung 404, Taiwan; [email protected] (S.-Y.L.); [email protected] (W.-H.H.); [email protected] (C.-C.L.); [email protected] (C.-H.T.); [email protected] (C.-H.L.); [email protected] (C.-Y.H.) 2 Division of Nephrology and Kidney Institute, China Medical University Hospital, Taichung 404, Taiwan 3 Division of Pulmonary and Critical Care Medicine, China Medical University Hospital and China Medical University, Taichung 404, Taiwan 4 Department of Family Medicine, China Medical University Hospital, Taichung 404, Taiwan 5 Management Office for Health Data, China Medical University Hospital, Taichung 404, Taiwan; [email protected] 6 College of Medicine, China Medical University, Taichung 404, Taiwan 7 Department of Orthopedics, China Medical University Hospital, Taichung 404, Taiwan; [email protected] 8 Department of Orthopedics, St. Martin De Porres Hospital, Chiayi 600, Taiwan; [email protected] 9 Department of Nuclear Medicine, China Medical University Hospital, Taichung 404, Taiwan 10 Department of Bioinformatics and Medical Engineering, Asia University, Taichung 413, Taiwan * Correspondence: [email protected]; Tel.: +886-4-2205-2121 (ext. -

The Neuroanatomy of Female Pelvic Pain
Chapter 2 The Neuroanatomy of Female Pelvic Pain Frank H. Willard and Mark D. Schuenke Introduction The female pelvis is innervated through primary afferent fi bers that course in nerves related to both the somatic and autonomic nervous systems. The somatic pelvis includes the bony pelvis, its ligaments, and its surrounding skeletal muscle of the urogenital and anal triangles, whereas the visceral pelvis includes the endopelvic fascial lining of the levator ani and the organ systems that it surrounds such as the rectum, reproductive organs, and urinary bladder. Uncovering the origin of pelvic pain patterns created by the convergence of these two separate primary afferent fi ber systems – somatic and visceral – on common neuronal circuitry in the sacral and thoracolumbar spinal cord can be a very dif fi cult process. Diagnosing these blended somatovisceral pelvic pain patterns in the female is further complicated by the strong descending signals from the cerebrum and brainstem to the dorsal horn neurons that can signi fi cantly modulate the perception of pain. These descending systems are themselves signi fi cantly in fl uenced by both the physiological (such as hormonal) and psychological (such as emotional) states of the individual further distorting the intensity, quality, and localization of pain from the pelvis. The interpretation of pelvic pain patterns requires a sound knowledge of the innervation of somatic and visceral pelvic structures coupled with an understand- ing of the interactions occurring in the dorsal horn of the lower spinal cord as well as in the brainstem and forebrain. This review will examine the somatic and vis- ceral innervation of the major structures and organ systems in and around the female pelvis. -

Thoracic Anatomy Autonomic Nervous System
Thoracic anatomy Autonomic nervous system Thoracic Anatomy 3.G.1.1 James Mitchell (December 24, 2003) Autonomic nervous system Division by direction Visceral efferent Preganglionic myelinated, postganglionic unmyelinated Synapse in ganglia Visceral afferent Similar to somatic afferent Cell body in CNS, peripheral processes travel with autonomic and somatic fibres Division by outflow Sympathetic Thoracolumbar outflow: T1-L3 Synapse in sympathetic trunk ganglia or other ganglia near CNS Preganglionic cholinergic, postganglionic predominantly noradrenergic (also adrenergic, cholinergic sudomotor and purinergic) Parasympathetic Craniosacral outflow: III, VII, IX, X, S2-4 Synapse adjacent to end-organs Cranial nerve parasympathetic ganglia are traversed by other fibres but contain only parasympathetic synapses Parasympathetic anatomy III Edinger-Westphal nuclei → oculomotor n. → n. to inferior oblique → ciliary ganglion → short ciliary nn. → ciliary muscle and sphincter pupillae VII Superior salivatory nucleus → nervus intermedius → facial n. → chorda tympani → lingual n. → submandibular ganglion → submandibular and sublingual glands Geniculate ganglion → greater petrosal n. → pterygopalatine ganglion → zygomatic and lacrimal nerves to lacrimal gland and nasal and palatine branches to nasal mucosa XI Inferior salivatory nucleus → glossopharyngeal nerve → tympanic plexus → lesser petrosal n. → otic ganglion → auriculotemporal n. → parotid gland and oral mucosa X Dorsal nucleus of vagus → vagus n. → minute ganglia in respiratory tract, heart, -

Sympathetic Tales: Subdivisons of the Autonomic Nervous System and the Impact of Developmental Studies Uwe Ernsberger* and Hermann Rohrer
Ernsberger and Rohrer Neural Development (2018) 13:20 https://doi.org/10.1186/s13064-018-0117-6 REVIEW Open Access Sympathetic tales: subdivisons of the autonomic nervous system and the impact of developmental studies Uwe Ernsberger* and Hermann Rohrer Abstract Remarkable progress in a range of biomedical disciplines has promoted the understanding of the cellular components of the autonomic nervous system and their differentiation during development to a critical level. Characterization of the gene expression fingerprints of individual neurons and identification of the key regulators of autonomic neuron differentiation enables us to comprehend the development of different sets of autonomic neurons. Their individual functional properties emerge as a consequence of differential gene expression initiated by the action of specific developmental regulators. In this review, we delineate the anatomical and physiological observations that led to the subdivision into sympathetic and parasympathetic domains and analyze how the recent molecular insights melt into and challenge the classical description of the autonomic nervous system. Keywords: Sympathetic, Parasympathetic, Transcription factor, Preganglionic, Postganglionic, Autonomic nervous system, Sacral, Pelvic ganglion, Heart Background interplay of nervous and hormonal control in particular The “great sympathetic”... “was the principal means of mediated by the sympathetic nervous system and the ad- bringing about the sympathies of the body”. With these renal gland in adapting the internal -
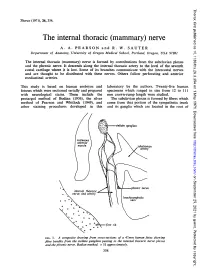
The Internal Thoracic (Mammary) Nerve A
Thorax: first published as 10.1136/thx.26.3.354 on 1 May 1971. Downloaded from Thorax (1971), 26, 354. The internal thoracic (mammary) nerve A. A. PEARSON and R. W. SAUTER Department of Anatomy, University of Oregon Medical School, Portland, Oregon, USA 97201 The internal thoracic (mammary) nerve is formed by contributions from the subclavian plexus and the phrenic nerve. It descends along the internal thoracic artery to the level of the seventh costal cartilage where it is lost. Some of its branches communicate with the intercostal nerves -and are thought to be distributed with these nerves. Others follow perforating and anterior mediastinal arteries. This study is based on human embryos and laboratory by the authors. Twenty-five human fetuses which were sectioned serially and prepared specimens which ranged in size from 12 to 111 with neurological stains. These include the mm crown-rump length were studied. protargol method of Bodian (1936), the silver The subclavian plexus is formed by fibres which method of Pearson and Whitlock (1949), and come from that portion of the sympathetic trunk other staining procedures developed in this and its ganglia which are located in the root of http://thorax.bmj.com/ on September 25, 2021 by guest. Protected copyright. FIG. 1. A composite drawing from cross-sections of a 47mm human fetus showing fibre bundles from the stellate ganglion passing to the internal thoracic nerve plexus and the phrenic nerve. Bodian method. x 51 approximately. 354 Thorax: first published as 10.1136/thx.26.3.354 on 1 May 1971. Downloaded from FIG. -

CVM 6100 Veterinary Gross Anatomy
2010 CVM 6100 Veterinary Gross Anatomy General Anatomy & Carnivore Anatomy Lecture Notes by Thomas F. Fletcher, DVM, PhD and Christina E. Clarkson, DVM, PhD 1 CONTENTS Connective Tissue Structures ........................................3 Osteology .........................................................................5 Arthrology .......................................................................7 Myology .........................................................................10 Biomechanics and Locomotion....................................12 Serous Membranes and Cavities .................................15 Formation of Serous Cavities ......................................17 Nervous System.............................................................19 Autonomic Nervous System .........................................23 Abdominal Viscera .......................................................27 Pelvis, Perineum and Micturition ...............................32 Female Genitalia ...........................................................35 Male Genitalia...............................................................37 Head Features (Lectures 1 and 2) ...............................40 Cranial Nerves ..............................................................44 Connective Tissue Structures Histologic types of connective tissue (c.t.): 1] Loose areolar c.t. — low fiber density, contains spaces that can be filled with fat or fluid (edema) [found: throughout body, under skin as superficial fascia and in many places as deep fascia] -

Development of the Rat Superior Cervical Ganglion: Initial Stages of Synapse Formation’
0270.6474/0503-0697$02.00/O The Journal of Neuroscience Copyright 0 Society for Neuroscience Vol. 5. No. 3, pp. 697-704 Printed in U.S.A. March 1985 Development of the Rat Superior Cervical Ganglion: Initial Stages of Synapse Formation’ ERIC RUBIN* Department of Physiology and Biophysics, Washington University School of Medicine, St. Louis, Missouri 63110 Abstract some aspects of synaptic organization through the prenatal period. Some of these results have been briefly reported (Rubin, 1982). Synapse formation in the rat superior cervical ganglion has been investigated electrophysiologically and at the ultra- Materials and Methods structural level. Preganglionic axons first enter the superior The procedures for obtaining and isolating fetal rats have been described cervical ganglion between days 12 and 13 of gestation (El2 (Rubin 1985a, b). As in the previous papers, the day of conception is counted to E13), and on El3 a postganglionic response can be evoked as embryonic day zero (EO), and the first postnatal day is termed PO. by preganglionic stimulation. The susceptibility of this re- flectrophysiology. In isolated fetuses, the right superior cervical ganglion sponse to fatigue and to blocking agents indicates that it is was exposed, along with the internal carotid nerve and the cervical sympa- mediated by cholinergic synapses. On E14, the overall thetic trunk (see Rubin, 1985a). The dissection was carried out at room strength of ganglionic innervation arising from different temperature in a standard Ringer’s solution (pH 7.2) of the following com- spinal segments already varies in a pattern resembling that position (in millimolar concentration): NaCI, 137.0; KCI, 4.0; MgCIP, 1.0; found in maturity. -

Sympathetic Control of Lower Esophageal Sphincter Function in the Cat: ACTION of DIRECT CERVICAL and SPLANCHNIC NERVE STIMULATION
Sympathetic Control of Lower Esophageal Sphincter Function in the Cat: ACTION OF DIRECT CERVICAL AND SPLANCHNIC NERVE STIMULATION Jacques Fournet, … , William J. Snape Jr., Sidney Cohen J Clin Invest. 1979;63(4):562-570. https://doi.org/10.1172/JCI109337. The purpose of this study was to determine the effect of direct stimulation of the sympathetic nerves on the lower esophageal sphincter (LES) in the anesthetized cat. Neither unilateral nor bilateral cervical sympathectomy, or splanchnicectomy significantly modified basal LES pressure in animals with intact vagi, or animals having undergone bilateral cervical vagotomy. Electrical stimulation of the cut, peripheral, cervical sympathetic trunk increased mean arterial blood pressure, but had no effect on LES pressure or LES relaxation as induced by vagal stimulation. Stimulation of the central end of the cervical sympathetic trunk had no effect on LES pressure. Stimulation of the central end of the cut splanchnic nerve produced a decrease in LES pressure with a maximal response of 69.1±16.0% (mean±SEM). This inhibitory response was not modified by either propranolol or bilateral cervical vagotomy. Stimulation of the peripheral end of the cut, greater splanchnic nerve gave an increase in LES pressure with a maximal response of 38.2±7.19 mm Hg. Guanethidine, in the presence or absence of the adrenal glands, significantly augmented this excitatory response. This response was also slightly increased by phentolamine alone at 10 V, 1 Hz, but was not altered by propranolol. The excitatory response was completely antagonized by atropine or by trimethaphan camsylate. Stimulation of the peripheral end of the splanchnic nerve inhibited LES relaxation as induced by […] Find the latest version: https://jci.me/109337/pdf Sympathetic Control of Lower Esophageal Sphincter Function in the Cat ACTION OF DIRECT CERVICAL AND SPLANCHNIC NERVE STIMULATION JACQUES FOURNET, WILLIAM J. -

Morphology of Sympathetic Chain in Saguinus Niger
Anais da Academia Brasileira de Ciências (2013) 85(1): 365-370 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 www.scielo.br/aabc Morphology of sympathetic chain in Saguinus niger MARINA P.E. PINTO1, ÉRIKA BRANCO1, EMERSON T. FIORETTO2, LUIZA C. PEREIRA3 and ANA R. LIMA1 1Universidade Federal Rural da Amazônia (UFRA), Instituto de Saúde e Produção Animal – ISPA, Faculdade de Medicina Veterinária, Avenida Perimetral, 2501, Belém, PA, Brasil 2 Universidade Federal de Sergipe (UFS), Cidade Universitária Professor José Aloísio de Campos, Avenida Marechal Rondon, s/n, Jardim Rosa Elze, São Cristovão, Aracajú, SE, Brasil 3 Empresa Hydro LTDA, Mina de Bauxita – Paragominas, PA, Brasil Manuscript received on March 20, 2012; accepted for publication on October 2, 2012 ABSTRACT Saguinus niger popularly known as Sauim, is a Brazilian North primate. Sympathetic chain investigation would support traumatic and/or cancer diagnosis which are little described in wild animals. The aim of this study was to describe the morphology and distribution of sympathetic chain in order to supply knowledge for neurocomparative research. Three female young animals that came death by natural causes were investigated. Animals were fixed in formaldehyde 10% and dissected along the sympathetic chain in neck, thorax and abdomen. Cranial cervical ganglion was located at the level of carotid bifurcation, related to carotid internal artery. In neck basis the vagosympathetic trunk divides into the sympathetic trunk and the parasympathetic vagal nerve. Sympathetic trunk ran in dorsal position and originated the stellate ganglia, formed by the fusion of caudal cervical and first thoracic ganglia. -

The Axatomy of the Autonomic Nervous System in the Dog1
THE AXATOMY OF THE AUTONOMIC NERVOUS SYSTEM IN THE DOG1 NICHOLAS JAMES AIIZERES Ucpartnitiit of Aiintoiiry, Cnzvemtty of Xtclizgaii Scliool of ;2/cdicmc. Ann Arbor, Mtclatgan ELEVEN FIGURES INTRODUCTTOX I<iiowledgc clerivccl from cmprimeiits on tlie clog has not infrequently been applied to man without considering dif- fcmnccs in aiiatoniy. This is c~spcciallytrue of the autonomic nervous systeni, oiily parts of which have b:mi described in the adult clog. The cardiac ncli'vcs \\'ere described by Sc1iuran.- Iew ( '27) ant1 Soniclez ( '39), the gcw~alplan of the abrloniirial ancl sacral regions by Trumble ( '34), tlic lunibosacral trunk by Alehler, Fisclier and Alexander ( '52), the vagi lsy Hilsa- beck aid Hill ( '50) and BIcC'rca and D'hrcy ( '%), ant1 the urogenital plexuses by Schal~adasch( '26) aiicl Ncdowar ( '2.3. The present study was undertaken to fill gaps in previous clescriptions ancl to present an over-all view of the autonomic iierrous system in the (log, esclutliiig the cephalic region. In the pi-c~seiitiiivcstigation the XI< terminology has been used iiistpad of the usual RNA familiar in human anatomy loccause tlic study was based on the clog. Sincc a dog is a pronograde niamnial the terms cranial aiid caudal seeni nior(t appropriat r tliaii the ESA terms superior and inferior. Tiiis paper is n condensation of a clissei t:ition snbniittctl in paytial fulfillmc.~it of tlrc rcqiiireiiieuts for the drgrce of Doctor of Pliilosopliy in the University of Rlicliigaii. I wish to tlimik Dr. It. T. Wootlhuine for Ills interest and aclriec. For the supply of nlatciial tlie author vihlies to e\-pr('\s his appreciatioir to mcmbc~is of the 1)ep:irtiiiciits of P1iTsiolog.y :iiiil PIiar~u:~cologrof tlic Viiirersity of hliclrigan.