The Intrinsic Cardiac Nervous System and Its Role in Cardiac Pacemaking and Conduction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Communication Center
2/16/2012 Communication Center Nervous System Regulation •The Neural System is only 3% of your body weight, but is the most complex organ system. •Nervous impulses are fast acting (milliseconds) Neural vs. Hormonal but short lived. Overview •The nervous system includes all neural tissue in the body. Basic units are: Two Anatomical Divisions of The Nervous System a. Neurons (individual nerve cells) 1. CNS: Central Nervous System b. Neuroglia • supporting cells • Brain & Spinal Cord • separate & protect the neurons • Responsible for integrating, processing, & • provide supporting framework coordinatinggy sensory data and motor commands. i.e.- stumble example • act as phagocytes • regulate composition of interstitial fluid • The brain is also the organ responsible for • a.k.a. glial cells intelligence, memory, learning, & emotion • outnumber neurons 1 2/16/2012 2. PNS: Peripheral Nervous System PNS has 2 functional divisions • All nervous tissue outside CNS Afferent Division (Sensory) • Carries sensory data to CNS, carries motor •Bring sensory information to CNS from receptors in commands from the CNS. peripheral nervous tissue & organs. • Bundles of nerve fibers carry impulses in the PNS are known as per ip hera l nerves or jus t “nerves”. Efferent Division (Motor) •Carries motor commands from CNS to muscles & • Nerves attached to the brain are called cranial glands, these target organs are called effectors. nerves. Nerves attached to the spinal cord are called spinal nerves. The Efferent Division is broken into Somatic & The ANS has a: Autonomic Components (SNS) Somatic System: controls skeletal muscle contractions these can be voluntary (conscious) or sympathetic division involuntary (unconscious) {reflexes}. } antagonistic effects parasympathetic (ANS) Au tonom ic Sys tem: * a.k.a. -

Cardiac Sympathectomy and Spinal Cord Stimulation Attenuate Reflex-Mediated Norepinephrine Release During Ischemia Preventing Ventricular Fibrillation
Cardiac sympathectomy and spinal cord stimulation attenuate reflex-mediated norepinephrine release during ischemia preventing ventricular fibrillation Jeffrey L. Ardell, … , J. Andrew Armour, Kalyanam Shivkumar JCI Insight. 2019. https://doi.org/10.1172/jci.insight.131648. Research In-Press Preview Cardiology Neuroscience Rationale: Reflex-mediated sympathoexcitation is central to the pathogenesis of arrhythmias and heart disease; neuraxial modulation can favorably attenuate these cardiac reflexes leading to cardioprotection. Objective: The purpose of this study was to define the mechanism by which cardiac neural decentralization and spinal cord stimulation (SCS) reduces ischemia-induced ventricular fibrillation (VF) and sudden cardiac death (SCD) by utilizing direct neurotransmitter measurements in the heart. Methods and Results: Direct measurement of norepinephrine (NE) levels in the left ventricular (LV) interstitial fluid (ISF) by microdialysis in response to transient left anterior descending coronary artery occlusion (CAO: 15 min) in anesthetized canines. Responses were studied with: (i) intact neuraxis and were compared to those in which the (ii) intrathoracic component of the cardiac neuraxis (stellate ganglia),(iii) the intrinsic cardiac neuronal (ICN) system were surgically delinked from the central nervous system versus (iv) subjects with intact neuraxis subjected to pre-emptive SCS (T1-T3 spinal level). With an intact neuraxis, animals with exaggerated NE-ISF levels in response to CAO were at increased risk for VF and SCD. During CAO there was a 152% increase in NE level when the entire neuraxis was intact compared to 114% following intrathoracic neuraxial decentralization (removal of the stellates) and 16% increase following ICN decentralization, when the entire heart and ICN was delinked from the other levels of the neuraxis. -

Autonomic Nerve Activity and Cardiac Arrhythmias
ACORP Complete (with appendices) Last Name of PI► Protocol No. Assigned by the IACUC►02002 Official Date of Approval► 1. Caging needs. Complete the table below to describe the housing that will have to be accommodated by the housing sites for this protocol: d. Is this housing e. Estimated c. Number of consistent with the maximum number a. Species b. Type of housing* individuals per Guide and USDA of housing units housing unit** regulations? needed at any one (yes/no***) time Chain link run, 3x6 Canines 1 no 7 and 4x10 feet cage *See ACORP Instructions, for guidance on describing the type of housing needed. If animals are to be housed according to a local Standard Operating Procedure (SOP), enter “standard (see SOP)” here, and enter the SOP into the table in Item Y. If the local standard housing is not described in a SOP, enter “standard, see below” in the table and describe the standard housing here: Chain link run, 3x6 feet cages ** The Guide states that social animals should generally be housed in stable pairs or groups. Provide a justification if any animals will be housed singly (if species is not considered “social”, then so note) Dogs are housed singly in chain link runs but can socialize with one another since each room has two to five dog runs. In addition, while their runs are being cleaned on a daily basis, pairs of dogs are allowed to exercise and play together in a designated "romper room". Animals are fitted with DSI transmitters and need to be housed singly in a cage for which DSI receivers are installed to receive signals from the transmitters. -

Clinical Presentations of Lumbar Disc Degeneration and Lumbosacral Nerve Lesions
Hindawi International Journal of Rheumatology Volume 2020, Article ID 2919625, 13 pages https://doi.org/10.1155/2020/2919625 Review Article Clinical Presentations of Lumbar Disc Degeneration and Lumbosacral Nerve Lesions Worku Abie Liyew Biomedical Science Department, School of Medicine, Debre Markos University, Debre Markos, Ethiopia Correspondence should be addressed to Worku Abie Liyew; [email protected] Received 25 April 2020; Revised 26 June 2020; Accepted 13 July 2020; Published 29 August 2020 Academic Editor: Bruce M. Rothschild Copyright © 2020 Worku Abie Liyew. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Lumbar disc degeneration is defined as the wear and tear of lumbar intervertebral disc, and it is mainly occurring at L3-L4 and L4-S1 vertebrae. Lumbar disc degeneration may lead to disc bulging, osteophytes, loss of disc space, and compression and irritation of the adjacent nerve root. Clinical presentations associated with lumbar disc degeneration and lumbosacral nerve lesion are discogenic pain, radical pain, muscular weakness, and cutaneous. Discogenic pain is usually felt in the lumbar region, or sometimes, it may feel in the buttocks, down to the upper thighs, and it is typically presented with sudden forced flexion and/or rotational moment. Radical pain, muscular weakness, and sensory defects associated with lumbosacral nerve lesions are distributed on -

'Voodoo' Death Revisited: the Modern Lessons of Neurocardiology†
KEYNOTE ADDRESS MARTIN A. SAMUELS, MD, DSc (hon), FAAN, MACP* Neurologist-in-Chief and Chairman Department of Neurology Brigham and Women’s Hospital Professor of Neurology Harvard Medical School Boston, MA ‘Voodoo’ death revisited: The modern lessons of neurocardiology† n 1942, Walter Bradford Cannon published a personal danger or threat of injury; (7) after danger is remarkable paper entitled “‘Voodoo’ Death,”1 in over; and (8) reunion, triumph, or happy ending. which he recounted anecdotal experiences, largely Common to all is that they involve events impossible I from the anthropology literature, of death from for the victim to ignore and to which the response is fright. These events, drawn from widely disparate parts overwhelming excitation, giving up, or both. of the world, had several features in common. They were In 1957, Carl Richter reported on a series of experi- all induced by an absolute belief that an external force, ments aimed at elucidating the mechanism of such as a wizard or medicine man, could, at will, cause Cannon’s “voodoo” death.3 Richter studied the length demise and that the victim himself had no power to alter of time domesticated rats could swim at various water this course. This perceived lack of control over a power- temperatures and found that at a water temperature of ful external force is the sine qua non for all the cases 93º these rats could swim for 60 to 80 minutes. recounted by Cannon, who postulated that death was However, if the animal’s whiskers were trimmed, it caused “by a lasting and intense action of the sym- would invariably drown within a few minutes. -

Simple Ways to Dissect Ciliary Ganglion for Orbital Anatomical Education
OkajimasDetection Folia Anat. of ciliary Jpn., ganglion94(3): 119–124, for orbit November, anatomy 2017119 Simple ways to dissect ciliary ganglion for orbital anatomical education By Ming ZHOU, Ryoji SUZUKI, Hideo AKASHI, Akimitsu ISHIZAWA, Yoshinori KANATSU, Kodai FUNAKOSHI, Hiroshi ABE Department of Anatomy, Akita University Graduate School of Medicine, Akita, 010-8543 Japan –Received for Publication, September 21, 2017– Key Words: ciliary ganglion, orbit, human anatomy, anatomical education Summary: In the case of anatomical dissection as part of medical education, it is difficult for medical students to find the ciliary ganglion (CG) since it is small and located deeply in the orbit between the optic nerve and the lateral rectus muscle and embedded in the orbital fat. Here, we would like to introduce simple ways to find the CG by 1): tracing the sensory and parasympathetic roots to find the CG from the superior direction above the orbit, 2): transecting and retracting the lateral rectus muscle to visualize the CG from the lateral direction of the orbit, and 3): taking out whole orbital structures first and dissecting to observe the CG. The advantages and disadvantages of these methods are discussed from the standpoint of decreased laboratory time and students as beginners at orbital anatomy. Introduction dissection course for the first time and with limited time. In addition, there are few clear pictures in anatomical The ciliary ganglion (CG) is one of the four para- textbooks showing the morphology of the CG. There are sympathetic ganglia in the head and neck region located some scientific articles concerning how to visualize the behind the eyeball between the optic nerve and the lateral CG, but they are mostly based on the clinical approaches rectus muscle in the apex of the orbit (Siessere et al., rather than based on the anatomical procedure for medical 2008). -

P2 Receptor-Mediated Modulation of Neurotransmitter Release—An Update
Purinergic Signalling (2007) 3:269–284 DOI 10.1007/s11302-007-9080-0 ORIGINAL ARTICLE P2 receptor-mediated modulation of neurotransmitter release—an update Beáta Sperlágh & Attila Heinrich & Cecilia Csölle Received: 13 July 2007 /Accepted: 28 August 2007 / Published online: 9 October 2007 # Springer Science + Business Media B.V. 2007 Abstract Presynaptic nerve terminals are equipped with a ENPP ectonucleotide pyrophosphatase number of presynaptic auto- and heteroreceptors, including EJP excitatory junction potential ionotropic P2X and metabotropic P2Y receptors. P2 recep- ENTPDase ectonucleoside triphosphate diphosphohydro- tors serve as modulation sites of transmitter release by ATP lase and other nucleotides released by neuronal activity and EPP end plate potential pathological signals. A wide variety of P2X and P2Y EPSC excitatory postsynaptic current receptors expressed at pre- and postsynaptic sites as well as EPSP excitatory postsynaptic potential in glial cells are involved directly or indirectly in the GABA +-aminobutyric acid modulation of neurotransmitter release. Nucleotides are GPCR G-protein coupled receptor released from synaptic and nonsynaptic sites throughout the IL-1β interleukin-1β nervous system and might reach concentrations high enough IPSC inhibitory postsynaptic current to activate these receptors. By providing a fine-tuning LC locus coeruleus mechanism these receptors also offer attractive sites for LPS lipopolysaccharide pharmacotherapy in nervous system diseases. Here we mEPP miniature EPP review the rapidly emerging data on the modulation of mEPSC miniature EPSC transmitter release by facilitatory and inhibitory P2 receptors NA noradrenaline and the receptor subtypes involved in these interactions. NMJ neuromuscular junction NT neurotransmitter Keywords Neuromodulation . Presynaptic . P2X receptor . NTS nucleus tractus solitarii P2Y receptors . -
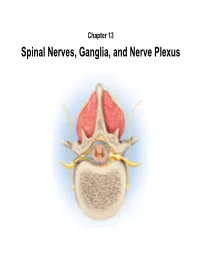
Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves
Chapter 13 Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves Posterior Spinous process of vertebra Posterior root Deep muscles of back Posterior ramus Spinal cord Transverse process of vertebra Posterior root ganglion Spinal nerve Anterior ramus Meningeal branch Communicating rami Anterior root Vertebral body Sympathetic ganglion Anterior General Anatomy of Nerves and Ganglia • Spinal cord communicates with the rest of the body by way of spinal nerves • nerve = a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue – mixed nerves contain both afferent (sensory) and efferent (motor) fibers – composed of thousands of fibers carrying currents in opposite directions Anatomy of a Nerve Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Epineurium Perineurium Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Endoneurium Nerve Rootlets fiber Posterior root Fascicle Posterior root ganglion Anterior Blood root vessels Spinal nerve (b) Copyright by R.G. Kessel and R.H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, 1979, W.H. Freeman, All rights reserved Blood vessels Fascicle Epineurium Perineurium Unmyelinated nerve fibers Myelinated nerve fibers (a) Endoneurium Myelin General Anatomy of Nerves and Ganglia • nerves of peripheral nervous system are ensheathed in Schwann cells – forms neurilemma and often a myelin sheath around the axon – external to neurilemma, each fiber is surrounded by -

Early Neuronal and Glial Fate Restriction of Embryonic Neural Stem Cells
The Journal of Neuroscience, March 5, 2008 • 28(10):2551–2562 • 2551 Development/Plasticity/Repair Early Neuronal and Glial Fate Restriction of Embryonic Neural Stem Cells Delphine Delaunay,1,2 Katharina Heydon,1,2 Ana Cumano,3 Markus H. Schwab,4 Jean-Le´on Thomas,1,2 Ueli Suter,5 Klaus-Armin Nave,4 Bernard Zalc,1,2 and Nathalie Spassky1,2 1Inserm, Unite´ 711, 75013 Paris, France, 2Institut Fe´de´ratif de Recherche 70, Faculte´deMe´decine, Universite´ Pierre et Marie Curie, 75013 Paris, France, 3Inserm, Unite´ 668, Institut Pasteur, 75724 Paris Cedex 15, France, 4Max-Planck-Institute of Experimental Medicine, D-37075 Goettingen, Germany, and 5Institute of Cell Biology, Swiss Federal Institute of Technology (ETH), ETH Ho¨nggerberg, CH-8093 Zu¨rich, Switzerland The question of how neurons and glial cells are generated during the development of the CNS has over time led to two alternative models: either neuroepithelial cells are capable of giving rise to neurons first and to glial cells at a later stage (switching model), or they are intrinsically committed to generate one or the other (segregating model). Using the developing diencephalon as a model and by selecting a subpopulation of ventricular cells, we analyzed both in vitro, using clonal analysis, and in vivo, using inducible Cre/loxP fate mapping, the fate of neuroepithelial and radial glial cells generated at different time points during embryonic development. We found that, during neurogenic periods [embryonic day 9.5 (E9.5) to 12.5], proteolipid protein ( plp)-expressing cells were lineage-restricted neuronal precursors, but later in embryogenesis, during gliogenic periods (E13.5 to early postnatal), plp-expressing cells were lineage-restricted glial precursors. -

GLOSSARY of MEDICAL and ANATOMICAL TERMS
GLOSSARY of MEDICAL and ANATOMICAL TERMS Abbreviations: • A. Arabic • abb. = abbreviation • c. circa = about • F. French • adj. adjective • G. Greek • Ge. German • cf. compare • L. Latin • dim. = diminutive • OF. Old French • ( ) plural form in brackets A-band abb. of anisotropic band G. anisos = unequal + tropos = turning; meaning having not equal properties in every direction; transverse bands in living skeletal muscle which rotate the plane of polarised light, cf. I-band. Abbé, Ernst. 1840-1905. German physicist; mathematical analysis of optics as a basis for constructing better microscopes; devised oil immersion lens; Abbé condenser. absorption L. absorbere = to suck up. acervulus L. = sand, gritty; brain sand (cf. psammoma body). acetylcholine an ester of choline found in many tissue, synapses & neuromuscular junctions, where it is a neural transmitter. acetylcholinesterase enzyme at motor end-plate responsible for rapid destruction of acetylcholine, a neurotransmitter. acidophilic adj. L. acidus = sour + G. philein = to love; affinity for an acidic dye, such as eosin staining cytoplasmic proteins. acinus (-i) L. = a juicy berry, a grape; applied to small, rounded terminal secretory units of compound exocrine glands that have a small lumen (adj. acinar). acrosome G. akron = extremity + soma = body; head of spermatozoon. actin polymer protein filament found in the intracellular cytoskeleton, particularly in the thin (I-) bands of striated muscle. adenohypophysis G. ade = an acorn + hypophyses = an undergrowth; anterior lobe of hypophysis (cf. pituitary). adenoid G. " + -oeides = in form of; in the form of a gland, glandular; the pharyngeal tonsil. adipocyte L. adeps = fat (of an animal) + G. kytos = a container; cells responsible for storage and metabolism of lipids, found in white fat and brown fat. -

The Neuroanatomy of Female Pelvic Pain
Chapter 2 The Neuroanatomy of Female Pelvic Pain Frank H. Willard and Mark D. Schuenke Introduction The female pelvis is innervated through primary afferent fi bers that course in nerves related to both the somatic and autonomic nervous systems. The somatic pelvis includes the bony pelvis, its ligaments, and its surrounding skeletal muscle of the urogenital and anal triangles, whereas the visceral pelvis includes the endopelvic fascial lining of the levator ani and the organ systems that it surrounds such as the rectum, reproductive organs, and urinary bladder. Uncovering the origin of pelvic pain patterns created by the convergence of these two separate primary afferent fi ber systems – somatic and visceral – on common neuronal circuitry in the sacral and thoracolumbar spinal cord can be a very dif fi cult process. Diagnosing these blended somatovisceral pelvic pain patterns in the female is further complicated by the strong descending signals from the cerebrum and brainstem to the dorsal horn neurons that can signi fi cantly modulate the perception of pain. These descending systems are themselves signi fi cantly in fl uenced by both the physiological (such as hormonal) and psychological (such as emotional) states of the individual further distorting the intensity, quality, and localization of pain from the pelvis. The interpretation of pelvic pain patterns requires a sound knowledge of the innervation of somatic and visceral pelvic structures coupled with an understand- ing of the interactions occurring in the dorsal horn of the lower spinal cord as well as in the brainstem and forebrain. This review will examine the somatic and vis- ceral innervation of the major structures and organ systems in and around the female pelvis. -

A Step Towards Stereotactic Navigation During Pelvic Surgery: 3D Nerve Topography
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Erasmus University Digital Repository Surgical Endoscopy and Other Interventional Techniques https://doi.org/10.1007/s00464-018-6086-3 A step towards stereotactic navigation during pelvic surgery: 3D nerve topography A. R. Wijsmuller1,2 · C. Giraudeau3 · J. Leroy4 · G. J. Kleinrensink5 · E. Rociu6 · L. G. Romagnolo7 · A. G. F. Melani7,8,9 · V. Agnus2 · M. Diana3 · L. Soler3 · B. Dallemagne2 · J. Marescaux2 · D. Mutter2 Received: 10 May 2017 / Accepted: 1 February 2018 © The Author(s) 2018. This article is an open access publication Abstract Background Long-term morbidity after multimodal treatment for rectal cancer is suggested to be mainly made up by nerve- injury-related dysfunctions. Stereotactic navigation for rectal surgery was shown to be feasible and will be facilitated by highlighting structures at risk of iatrogenic damage. The aim of this study was to investigate the ability to make a 3D map of the pelvic nerves with magnetic resonance imaging (MRI). Methods A systematic review was performed to identify a main positional reference for each pelvic nerve and plexus. The nerves were manually delineated in 20 volunteers who were scanned with a 3-T MRI. The nerve identifiability rate and the likelihood of nerve identification correctness were determined. Results The analysis included 61 studies on pelvic nerve anatomy. A main positional reference was defined for each nerve. On MRI, the sacral nerves, the lumbosacral plexus, and the obturator nerve could be identified bilaterally in all volunteers. The sympathetic trunk could be identified in 19 of 20 volunteers bilaterally (95%).