Pathogen-Free, Plasma-Poor Platelet Lysate and Expansion of Human Mesenchymal Stem Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Blood, Lymph, and Immunity Joann Colville
Blood, Lymph, and Immunity Joann Colville CHAPTER OUTLINE .»: BLOOD Function Introduction Lymphatic Structures Plasma THE IMMUNE SYSTEM Cellular Components of Blood Function Red Blood Cells Immune Reactions Platelets Immunization: Protection Against Disease White Blood Cells THE LYMPHATICSYSTEM Lymph Formation Characteristics LEARNING OBJECTIVES • List and describe the functions of blood • List the functions of the lymphatic system • Describe the composition of blood plasma • Describe the structure and functions of the lymph nodes, • Describe the characteristics of mature erythrocytes spleen, thymus, tonsils, and GALT Describe the structure of the hemoglobin molecule and • List the functions of the immune system explain the fate of hemoglobin following intravascular and • Differentiate between specific and nonspecific immune extravascular hemolysis reactions • Give the origin of thrombocytes and describe their • Differentiate between cell-mediated and humoral immunity characteristics and functions • List the components involved in cell-mediated immunity • Listthe types of leukocytes and describe the functions of each and explain the role of each • Describe the formation of lymph fluid and its circulation List and describe the classes of immunoglobulins through the lymphatic system • Differentiate between active and passive immunity BLOOD vessels of the cardiovascular system. It has three main func- tions: transportation, regulation, and defense. INTRODUCTION Blood. Say the word, and some people cringe, others faint, Function and if you believe authors Bram Stoker, Stephen King, and 1. Blood is a transport system. Christopher Moore (all authors of vampire books), others • It carries oxygen, nutrients, and other essential com- drool. But no matter what you think about blood, animals pounds to every living cell in the body. -

The Diagnosis of Paroxysmal Nocturnal Hemoglobinuria: Role of Flow Cytometry
THE DIAGNOSIS OF PAROXYSMAL NOCTURNAL HEMOGLOBINURIA: ROLE OF FLOW CYTOMETRY Alberto Orfao Department of Medicine, Cancer Research Centre (IBMCC-CSIC/USAL), University of Salamanca, Salamanca, Spain Paroxysmal nocturnal hemoglobinuria (PNH) on one or multiple populations of peripheral blood is an acquired clonal disorder typically affecting red cells and/or leucocytes of PNH patients, this young adults, which involves the phosphatidylino- translating into a new diagnostic test. Because sitol glycan anchor biosynthesis class A gene of this flow cytometry became an essential tool (PIGA) coded in chromosome X, in virtually every in the diagnosis of PNH. In turn, it could be also case. The altered gene encodes for a defective pro- demonstrated that investigation of the presence of tein product, the pig-a enzyme which is involved GPI-deficient cells among circulating mature neu- in the early steps of the synthesis of glycosylphos- trophils and monocytes, increases the sensitivity phatidylinositol (GPI). This later molecule (GPI) of red blood cell screening because of the shorter acts as an anchor of a wide range of proteins to lifetime of both populations of leucocytes. Despite the cell surface. At present a large number of GPI- all the above, routine assessment of CD55 and anchor associated proteins have been described CD59 is also associated with several limitations which are expressed by one or multiple distinct due to distinct patterns of expression among dif- compartments of hematopoietic cells. The altered ferent cell populations, dim and heterogeneous PIGA gene leads to an altered GPI anchor which expression among normal individuals and the lack leads to defective expression of GPI-AP on the of experience in many labs as regards the normal cytoplasmic membrane, on the cell surface. -

Chapter 06 Lecture Outline
Chapter 06 Lecture Outline See separate PowerPoint slides for all figures and tables pre- inserted into PowerPoint without notes. Copyright © 2016 McGraw-Hill Education. Permission required for reproduction or display. 1 Cardiovascular System: Blood © SPL/Science Source, (inset) © Andrew Syred/Science Source 2 Points to ponder • What type of tissue is blood and what are its components? • What is found in plasma? • Name the three formed elements in blood and their functions. • How does the structure of red blood cells relate to their function? • Describe the structure and function of each white blood cell. • What are disorders of red blood cells, white blood cells, and platelets? • What do you need to know before donating blood? • What are antigens and antibodies? • How are ABO blood types determined? • What blood types are compatible for blood transfusions? • What is the Rh factor and how is this important to pregnancy? • How does the cardiovascular system interact with other systems to maintain homeostasis? 3 6.1 Blood: An Overview What are the functions of blood? • Transportation: oxygen, nutrients, wastes, carbon dioxide, and hormones • Defense: against invasion by pathogens • Regulatory functions: body temperature, water- salt balance, and body pH 4 6.1 Blood: An Overview What is the composition of blood? • Remember: blood is a fluid connective tissue. • Formed elements are produced in red bone marrow. – Red blood cells/erythrocytes (RBCs) – White blood cells/leukocytes (WBCs) – Platelets/thrombocytes 5 6.1 Blood: An Overview What is the composition of blood? • Plasma – It consists of 91% water and 9% salts (ions) and organic molecules. – Plasma proteins are the most abundant organic molecules. -
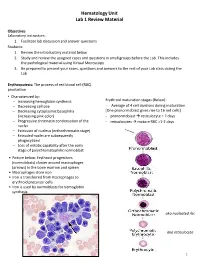
Hematology Unit Lab 1 Review Material
Hematology Unit Lab 1 Review Material Objectives Laboratory instructors: 1. Facilitate lab discussion and answer questions Students: 1. Review the introductory material below 2. Study and review the assigned cases and questions in small groups before the Lab. This includes the pathological material using Virtual Microscopy 3. Be prepared to present your cases, questions and answers to the rest of your Lab class during the Lab Erythropoiesis: The process of red blood cell (RBC) production • Characterized by: − Increasing hemoglobin synthesis Erythroid maturation stages (Below): − Decreasing cell size - Average of 4 cell divisions during maturation − Decreasing cytoplasmic basophilia [One pronormoblast gives rise to 16 red cells] (increasing pink color) - pronormoblast → reticulocyte = 7 days − Progressive chromatin condensation of the - reticulocytes → mature RBC =1-2 days nuclei − Extrusion of nucleus (orthochromatic stage) − Extruded nuclei are subsequently phagocytized − Loss of mitotic capability after the early stage of polychromatophilic normoblast • Picture below: Erythroid progenitors (normoblasts) cluster around macrophages (arrows) in the bone marrow and spleen • Macrophages store iron • Iron is transferred from macrophages to erythroid precursor cells • Iron is used by normoblasts for hemoglobin synthesis aka nucleated rbc aka reticulocyte 1 Mature Red Blood Cell 7-8 microns; round / ovoid biconcave disc with orange-red cytoplasm, no RNA, no nucleus; survives ~120 days in circulation Classification of Anemia by Morphology 1. -

CD59: a Long-Known Complement Inhibitor Has Advanced to a Blood Group System
B LOOD G ROUP R EVIEW CD59: A long-known complement inhibitor has advanced to a blood group system C. Weinstock, M. Anliker, and I. von Zabern The blood group system number 35 is based on CD59, a 20-kDa by Zalman et al.1 from the group of H.J. Muller-Eberhard in La membrane glycoprotein present on a large number of different Jolla, California, and in 1988 by Schönermark et al. from the cells, including erythrocytes. The major function of CD59 is to group of G.M. Hänsch in Heidelberg (Germany).2 This inhibitor protect cells from complement attack. CD59 binds to complement components C8 and C9 and prevents the polymerization of C9, was published under the designations “HRF (homologous which is required for the formation of the membrane attack restriction factor)” and “C8bp (C8 binding protein),” complex (MAC). Other functions of CD59 in cellular immunity respectively, to describe its properties.1,2 “C8bp” indicates the are less well defined. CD59 is inserted into the membrane by a glycosylphosphatidylinositol (GPI) anchor. A defect of this anchor binding capacity for C8. The name “HRF” points to a species causes lack of this protein from the cell membrane, which leads to incompatibility of this “factor” that provides protection from an enhanced sensitivity towards complement attack. Patients with complement attack more effectively in a homologous (e.g., paroxysmal nocturnal hemoglobinuria (PNH) harbor a varying human erythrocytes as a target of human complement) than in percentage of red blood cell clones with a defect in GPI-anchored proteins, including CD59. The most characteristic symptoms of a heterologous system (e.g., human erythrocytes as a target of this disease are episodes of hemolysis and thromboses. -

SEED Haematology Sysmex Educational Enhancement and Development October 2012
SEED Haematology Sysmex Educational Enhancement and Development October 2012 The red blood cell indices The full blood count has been used in conjunction with the traditional red The complete blood count (CBC) is central to clinical deci- cell indices in order to narrow down the possible causes sion making. This makes it one of the commonest laboratory of anaemia in an individual patient. investigations performed worldwide. Whilst the definition of what constitutes an CBC is influenced by the number Impedance technology and type of parameters measured by different haematology The RBC, HCT and MCV are all closely interrelated as they analysers, the traditional red cell indices that are widely are derived from information obtained from the passage used to classify anaemias are common to all. of cells through the aperture of the impedance channel of an automated haematology analyser. The impedance The laboratory approach to anaemia technology is based on the principle that an electrical field, Anaemia is an extremely common global healthcare prob- created between two electrodes of opposite charge, can lem. However, anaemia is merely a symptom which can be used to count and determine the size of cells. Blood result from a multitude of causes. Effective treatment is cells are poor conductors of electricity. The diluent in which only possible if the underlying cause is correctly identified. they are suspended as they pass through the aperture To this end, several classification systems have been devis- during counting is an isotonic solution which is a good ed. The most useful and widely used classification system conductor of electricity. -

Apheresis Red Cell Exchange/Transfusions
APHERESIS RED CELL EXCHANGE/TRANSFUSIONS In a patient treated in Manchester, parasitemia was virtually eliminated over eight hours by a 3.5 liter exchange blood transfusion (Plasmodium Falciparum Hyperparasitemia: Use of Exchange Transfusion in Seven Patients and a Review of the Literature). Several cases of severe babesiosis refractory to appropriate antibiotic therapy have been reported to respond promptly and dramatically to red blood cell (RBC) exchange transfusion. Asplenic patients, however, generally have a more severe course of illness, with hemolytic anemia, acute renal failure, disseminated intravascular coagulation, and pulmonary edema. Primary therapy is with antibiotics including clindamycin and quinine, with RBC exchange transfusion reported to be effective in severe cases. The RBC exchange transfusions succeeded in reducing significantly the level of parasitemia, dramatically improving the condition of an extremely ill patient. Our report adds to the small but growing literature on severe Babesia infection in humans, and provides further evidence to support the use of RBC exchange transfusion to treat severe babesiosis. Its single great advantage over antibiotic therapy is its rapid therapeutic effectiveness (Treatment of Babesiosis by Red Blood Cell Exchange in an HIV-Positive Splenectomized Patient). There was rapid clinical improvement after the whole-blood exchange transfusion. In cases of severe babesiosis, prompt institution of whole-blood exchange transfusion, in combination with appropriate antimicrobial therapy, can be life-saving. In patients with progressive babesiosis, early intervention with exchange transfusion, along with appropriate antimicrobial therapy, should be considered to speed clinical recovery. (Fulminant babesiosis treated with clindamycin, quinine, and whole-blood exchange transfusion. However, asplenic patients may have a much more serious clinical course. -

Indicators of the Iron Status of Populations: Red Blood Cell Parameters Sean Lynch
AnneX 1 Indicators of the iron status of populations: red blood cell parameters SEAN LYNch 19 Indicators OF the iron statUS OF popUlations: red blood cell parameters Contents 1. Introduction 22 2. Relationship between anaemia and iron deficiency 23 2.1 Physiological control of haemoglobin levels 23 2.2 Relationship between iron deficiency and anaemia 24 3. Red blood cell parameters 26 3.1 Haemoglobin 26 3.2 Hematocrit or packed cell volume 26 3.3 Mean cell volume and mean cell haemoglobin 27 3.4 Red cell distribution width 27 3.5 Reticulocyte haemoglobin concentration and percentage of hypochromic red cells 27 4. Role of haemoglobin as a screening indicator for iron deficiency 27 5. Conclusions 28 6. References 29 21 Assessing the iron statUS OF popUlations 1. Introduction Iron is an essential nutrient that plays a central role in many metabolic processes. Aerobic metabolism is critically dependent on maintaining normal concentrations of several iron-containing proteins that mediate oxygen transport, storage and utili- zation, particularly when the tissue demand for oxygen is increased by physical ac- tivity. Pioneering research over the last 50 years, much of it stemming from concepts developed and validated experimentally by Dr Clement Finch and his coworkers, led to the recognition that a negative iron balance resulting from an iron intake insuf- ficient to match losses from the body despite compensatory changes in the rate of absorption and, to a more limited extent, excretion could be divided into three stages based on the severity of the potential effect on physiological functions. The evalua- tion of functional impairment was related entirely to erythropoiesis for two reasons: the effects of changes in iron status on blood elements are readily evaluated, while the effect on the enzymes in other tissues necessitates obtaining biopsy samples. -

Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods
Guidance for Industry Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods Additional copies of this guidance document are available from: Office of Communication, Training and Manufacturers Assistance (HFM-40) 1401 Rockville Pike, Rockville, MD 20852-1448 (Tel) 1-800-835-4709 or 301-827-1800 (Internet) http://www.fda.gov/cber/guidelines.htm U.S. Department of Health and Human Services Food and Drug Administration Center for Biologics Evaluation and Research (CBER) January 2001 Technical Correction February 2001 TABLE OF CONTENTS Note: Page numbering may vary for documents distributed electronically. I. INTRODUCTION ............................................................................................................. 1 II. BACKGROUND................................................................................................................ 1 III. CHANGES FROM THE DRAFT GUIDANCE .............................................................. 2 IV. RECOMMENDED DONOR SELECTION CRITERIA FOR THE AUTOMATED RED BLOOD CELL COLLECTION PROTOCOLS ..................................................... 3 V. RECOMMENDED RED BLOOD CELL PRODUCT QUALITY CONTROL............ 5 VI. REGISTRATION AND LICENSING PROCEDURES FOR THE MANUFACTURE OF RED BLOOD CELLS COLLECTED BY AUTOMATED METHODS.................. 7 VII. ADDITIONAL REQUIREMENTS.................................................................................. 9 i GUIDANCE FOR INDUSTRY Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods This -

Package Insert
[US] 1 / 2 • Caution: Do not pipette by mouth. The absence of all viruses has not been determined. Blood Grouping Reagent • Caution: This Product Contains Natural Rubber Latex Which May Cause Allergic Reactions. Anti-K (KEL1) • Warning: Contains sodium azide (NaN3), which may react with lead or ® Seraclone Human Monoclonal copper plumbing to form explosive azides. If discarded in the sink, flush with (MS56) large amounts of water to prevent the build-up of explosive metal azides. • The bovine albumin used for the production of this reagent is sourced from FOR IN VITRO DIAGNOSTIC USE donor animals of U.S. origin that have been inspected and certified by U.S. For Tube Testing Veterinary Service inspectors to be disease free. MEETS FDA POTENCY REQUIREMENTS • Consult downloads.bio-rad.com to download the valid version of this instruction for use. U.S. License Number: 1845 Rx only SPECIMEN COLLECTION Fresh samples of clotted, EDTA or citrate anticoagulated whole blood collected PACKAGE SIZE following general blood sampling guidelines are acceptable. The specimen ® [REF] 808090100 [VOL] 5 mL Seraclone Anti-K (KEL1) should be tested as soon as possible after collection. If testing is delayed, specimens should be stored at 2 to 8°C, citrated specimens (donor segments) INTENDED USE may be stored at temperatures as low as 1°C. For the determination of the Kell (KEL1) antigen of red blood cells using the tube Note: Blood specimens exhibiting gross hemolysis or contamination should not test. be used. Clotted samples or those collected in EDTA may be tested within ten days from SUMMARY collection. -

And Frog Blood Cells
Human (Mammalian) and Frog Blood Cells Humans vs Frogs Human and frog are two kinds of animals with different evolutionary paths. Humans are mammals while frogs are amphibians. The three main types of blood cells are red blood cells, white blood cells, and platelets. The main difference between human blood cells and frog blood cells is that human red blood cells lack nuclei whereas frog blood cells contain nuclei. The white blood cells of both human and frog are similar to each other by means of morphology as well as function. Key Areas Covered 1. What are Human Blood Cells – Definition, Blood Cell Types, Function 2. What are Frog Blood Cells – Definition, Blood Cell Types, Function 3. What are the Similarities Between Human and Frog Blood Cells – Outline of Common Features 4. What is the Difference Between Human and Frog Blood Cells – Comparison of Key Differences Key Terms: Agranulocytes, Frog Blood Cells, Granulocytes, Human Blood Cells, Red Blood Cells, Red Cell Nuclei, Platelets What are Human Blood Cells Human blood cells are the circulating cells in the human blood. Human blood is composed of blood cells and plasma (the fluid component of blood). An adult human has about 5 liters of blood and blood cells take up about 2 liters of the total volume of the blood. The rest of the volume (~3 liters) consists of plasma. Blood cells are suspended in plasma. The three main types of blood cells in humans are red blood cells (erythrocytes), white blood cells (leukocytes), and platelets (thrombocytes). The most common type of blood cells are red blood cells. -

Blood and Immunity
Chapter Ten BLOOD AND IMMUNITY Chapter Contents 10 Pretest Clinical Aspects of Immunity Blood Chapter Review Immunity Case Studies Word Parts Pertaining to Blood and Immunity Crossword Puzzle Clinical Aspects of Blood Objectives After study of this chapter you should be able to: 1. Describe the composition of the blood plasma. 7. Identify and use roots pertaining to blood 2. Describe and give the functions of the three types of chemistry. blood cells. 8. List and describe the major disorders of the blood. 3. Label pictures of the blood cells. 9. List and describe the major disorders of the 4. Explain the basis of blood types. immune system. 5. Define immunity and list the possible sources of 10. Describe the major tests used to study blood. immunity. 11. Interpret abbreviations used in blood studies. 6. Identify and use roots and suffixes pertaining to the 12. Analyse several case studies involving the blood. blood and immunity. Pretest 1. The scientific name for red blood cells 5. Substances produced by immune cells that is . counteract microorganisms and other foreign 2. The scientific name for white blood cells materials are called . is . 6. A deficiency of hemoglobin results in the disorder 3. Platelets, or thrombocytes, are involved in called . 7. A neoplasm involving overgrowth of white blood 4. The white blood cells active in adaptive immunity cells is called . are the . 225 226 ♦ PART THREE / Body Systems Other 1% Proteins 8% Plasma 55% Water 91% Whole blood Leukocytes and platelets Formed 0.9% elements 45% Erythrocytes 10 99.1% Figure 10-1 Composition of whole blood.