Red Cell Agglutination in Non-Humans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Blood, Lymph, and Immunity Joann Colville
Blood, Lymph, and Immunity Joann Colville CHAPTER OUTLINE .»: BLOOD Function Introduction Lymphatic Structures Plasma THE IMMUNE SYSTEM Cellular Components of Blood Function Red Blood Cells Immune Reactions Platelets Immunization: Protection Against Disease White Blood Cells THE LYMPHATICSYSTEM Lymph Formation Characteristics LEARNING OBJECTIVES • List and describe the functions of blood • List the functions of the lymphatic system • Describe the composition of blood plasma • Describe the structure and functions of the lymph nodes, • Describe the characteristics of mature erythrocytes spleen, thymus, tonsils, and GALT Describe the structure of the hemoglobin molecule and • List the functions of the immune system explain the fate of hemoglobin following intravascular and • Differentiate between specific and nonspecific immune extravascular hemolysis reactions • Give the origin of thrombocytes and describe their • Differentiate between cell-mediated and humoral immunity characteristics and functions • List the components involved in cell-mediated immunity • Listthe types of leukocytes and describe the functions of each and explain the role of each • Describe the formation of lymph fluid and its circulation List and describe the classes of immunoglobulins through the lymphatic system • Differentiate between active and passive immunity BLOOD vessels of the cardiovascular system. It has three main func- tions: transportation, regulation, and defense. INTRODUCTION Blood. Say the word, and some people cringe, others faint, Function and if you believe authors Bram Stoker, Stephen King, and 1. Blood is a transport system. Christopher Moore (all authors of vampire books), others • It carries oxygen, nutrients, and other essential com- drool. But no matter what you think about blood, animals pounds to every living cell in the body. -

The Diagnosis of Paroxysmal Nocturnal Hemoglobinuria: Role of Flow Cytometry
THE DIAGNOSIS OF PAROXYSMAL NOCTURNAL HEMOGLOBINURIA: ROLE OF FLOW CYTOMETRY Alberto Orfao Department of Medicine, Cancer Research Centre (IBMCC-CSIC/USAL), University of Salamanca, Salamanca, Spain Paroxysmal nocturnal hemoglobinuria (PNH) on one or multiple populations of peripheral blood is an acquired clonal disorder typically affecting red cells and/or leucocytes of PNH patients, this young adults, which involves the phosphatidylino- translating into a new diagnostic test. Because sitol glycan anchor biosynthesis class A gene of this flow cytometry became an essential tool (PIGA) coded in chromosome X, in virtually every in the diagnosis of PNH. In turn, it could be also case. The altered gene encodes for a defective pro- demonstrated that investigation of the presence of tein product, the pig-a enzyme which is involved GPI-deficient cells among circulating mature neu- in the early steps of the synthesis of glycosylphos- trophils and monocytes, increases the sensitivity phatidylinositol (GPI). This later molecule (GPI) of red blood cell screening because of the shorter acts as an anchor of a wide range of proteins to lifetime of both populations of leucocytes. Despite the cell surface. At present a large number of GPI- all the above, routine assessment of CD55 and anchor associated proteins have been described CD59 is also associated with several limitations which are expressed by one or multiple distinct due to distinct patterns of expression among dif- compartments of hematopoietic cells. The altered ferent cell populations, dim and heterogeneous PIGA gene leads to an altered GPI anchor which expression among normal individuals and the lack leads to defective expression of GPI-AP on the of experience in many labs as regards the normal cytoplasmic membrane, on the cell surface. -

Chapter 06 Lecture Outline
Chapter 06 Lecture Outline See separate PowerPoint slides for all figures and tables pre- inserted into PowerPoint without notes. Copyright © 2016 McGraw-Hill Education. Permission required for reproduction or display. 1 Cardiovascular System: Blood © SPL/Science Source, (inset) © Andrew Syred/Science Source 2 Points to ponder • What type of tissue is blood and what are its components? • What is found in plasma? • Name the three formed elements in blood and their functions. • How does the structure of red blood cells relate to their function? • Describe the structure and function of each white blood cell. • What are disorders of red blood cells, white blood cells, and platelets? • What do you need to know before donating blood? • What are antigens and antibodies? • How are ABO blood types determined? • What blood types are compatible for blood transfusions? • What is the Rh factor and how is this important to pregnancy? • How does the cardiovascular system interact with other systems to maintain homeostasis? 3 6.1 Blood: An Overview What are the functions of blood? • Transportation: oxygen, nutrients, wastes, carbon dioxide, and hormones • Defense: against invasion by pathogens • Regulatory functions: body temperature, water- salt balance, and body pH 4 6.1 Blood: An Overview What is the composition of blood? • Remember: blood is a fluid connective tissue. • Formed elements are produced in red bone marrow. – Red blood cells/erythrocytes (RBCs) – White blood cells/leukocytes (WBCs) – Platelets/thrombocytes 5 6.1 Blood: An Overview What is the composition of blood? • Plasma – It consists of 91% water and 9% salts (ions) and organic molecules. – Plasma proteins are the most abundant organic molecules. -

Serotyping of Bdellovibrios by Agglutination and Indirect Immunofluorescence
INTERNATIONAL JOURNAL OF SYSTEMATICBACT~RIOLOGY. Oct. 1983, p. 816-821 Vol. 33, No. 4 0020-7713/83/040816-06$02.00/0 Copyright 0 1983, Internationnl Union of Microbiologicdl Societie\ Serotyping of Bdellovibrios by Agglutination and Indirect Immunofluorescence M. E. SCHELLING’* AND S. F. CONTI’ Department Biology, Texus A &M Unitwsit?, College Station, Texus 77843‘ und University of Mussa ch us et t s , A rn ii erst, Muss N c h iiset t J 0 1 002’ A comparative serological study was undertaken to clarify the uncertain interrelationships among various bdellovibrios. A total of 17 predacious (host- dependent) strains and 12 nonpredacious strains were serotyped. Strains were grouped into serotypes on the basis of positive agglutination and strongly positive fluorescence cross-reactions. The results of division of the strains into serogroups by these two methods were identical. Bdellovihrio starrii strain A3.12 and Bdellovibrio stolpii strain UKi2 were found to be antigenically distinct from each other and from the group of strains comprising Bdellovihrio hacteriovorus; the latter was found to be an antigenically heterogeneous group consisting of at least nine serogroups. All nonpredacious strains were found to be related antigenically to their obligately predacious counterparts. An antigen(s) common to all of the Bdellovibrio strains examined was exhibited as weakly positive fluorescence. Selected strains of Bdellovihrio were studied by Ouchterlony immunodiffusion, which confirmed the serogrouping results and provided a technically simple method of serotyping. Further immunochemical characterization of the strains of Bdellovihrio in conjunction with increased knowledge of other differences among strains will likely result in the formation of additional species of Bdellovihrio. -

Red Blood Cell Preparation and Hemagglutination Assay
Red Blood Cell Preparation and Hemagglutination Assay Purpose: This protocol describes the preparation of RBCs for storage and use in Hemagluttination assays, which are used to determine the La Sota B1 lentogenic Newcastle Disease virion hemaglutinin-neuraminidase titer relative to virion stock positive controls. The protocol was adapted from McGinnes et al. (2006) and “Detection of Hemagglutinating Viruses” (n.d.). Materials: ● Whole Blood Cells ● 50mL conical tube ● Multichannel micropipette, micropipette, and tips ● PBS with 2mM Penicillin/Streptomycin ● SV3 ● Alsever’s Solution ● 96 round bottom well plate ● Microscope slide ● Virion stock ● Centrifuge Procedure: 1. RBC preparation a. Obtain 25mL whole blood and add 25mL cold Alsever’s Solution for anti-coagulation in a 50mL conical tube and keep on ice b. Centrifuge whole blood at 500 RCF for 10min c. Aspirate blood plasma, buffy layer, and top erythrocytes d. Wash 3 times by resuspending in PBS with 2mM Penicillin/Streptomycin to double the pellet volume, centrifuging at 500 RCF, and aspirating the supernatant e. Resuspend the pellet in PBS with 2mM Penicillin/Streptomycin or SV3 for a final concentration of 10% pellet volume per total volume, and store at 2-7˚C for 1 week or 42 days 2. HA Titer a. Thaw your virion stock and samples on ice b. Use a multichannel micropipette to add 50µL of cold PBS to each row on a 96 round bottom well plate for each sample in duplicates c. Add 50µL of each sample to column 1 of their respective duplicate rows i. Don’t add anything to the negative control rows and add virion stocks to the positive control rows d. -

20 Hemolytic Anemias Due to Abnormal Red Cell Enzymes
Hemolytic Anemias Due to Abnormal Red Cell Enzymes MODULE Hematology and Blood Bank Technique 20 HEMOLYTIC ANEMIAS DUE TO Notes ABNORMAL RED CELL ENZYMES 20.1 INTRODUCTION The main metabolic substrate for the RBCs is glucose. It is metabolized by two pathways: approximately 90% of the glucose is metabolized through the Embden Meyerhoff (glycolytic) pathway and the rest by the hexose monophosphate (HMP) pathway. In the Embden Meyerhoff (glycolytic) pathway glucose is metabolized to lactate through a series of enzymatic steps. Each molecule of glucose gives rise to 2 molecules of ATP. The ATP provides energy to maintain red cell volume, shape and flexibility. An ATP dependent pump in the red cell membrane actively keeps sodium out of the cell and potassium inside. The red cell has the enzymes that are needed for the glycolytic pathway. These enzymes help break down glucose to generate ATP which is the source of energy. About 10% of the glucose is diverted to the Hexose Monophosphate shunt pathway and this is essential for protection of red cells from oxidative stress. This pathway is necessary for the generation of NADPH which then reduces oxidized glutathione (GSSG) to reduced glutathione (GSH). GSH prevents the accumulation of H2O2 and the oxidation of hemoglobin to methemoglobin. When the level of GSH falls, H2O2 accumulates in the cell and oxidizes the hemoglobin to methemoglobin which becomes denatured and precipitates as Heinz bodies. These inclusions are rigid and attached to the red cell membrane and make the red cell susceptible to hemolysis. The NADPH required in this pathway is generated by the enzyme Glucose 6 phosphate dehydrogenase (G6PD). -
Hematology Unit Lab 1 Review Material
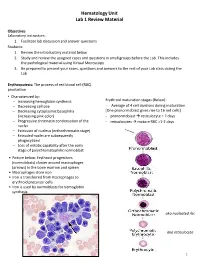
Hematology Unit Lab 1 Review Material Objectives Laboratory instructors: 1. Facilitate lab discussion and answer questions Students: 1. Review the introductory material below 2. Study and review the assigned cases and questions in small groups before the Lab. This includes the pathological material using Virtual Microscopy 3. Be prepared to present your cases, questions and answers to the rest of your Lab class during the Lab Erythropoiesis: The process of red blood cell (RBC) production • Characterized by: − Increasing hemoglobin synthesis Erythroid maturation stages (Below): − Decreasing cell size - Average of 4 cell divisions during maturation − Decreasing cytoplasmic basophilia [One pronormoblast gives rise to 16 red cells] (increasing pink color) - pronormoblast → reticulocyte = 7 days − Progressive chromatin condensation of the - reticulocytes → mature RBC =1-2 days nuclei − Extrusion of nucleus (orthochromatic stage) − Extruded nuclei are subsequently phagocytized − Loss of mitotic capability after the early stage of polychromatophilic normoblast • Picture below: Erythroid progenitors (normoblasts) cluster around macrophages (arrows) in the bone marrow and spleen • Macrophages store iron • Iron is transferred from macrophages to erythroid precursor cells • Iron is used by normoblasts for hemoglobin synthesis aka nucleated rbc aka reticulocyte 1 Mature Red Blood Cell 7-8 microns; round / ovoid biconcave disc with orange-red cytoplasm, no RNA, no nucleus; survives ~120 days in circulation Classification of Anemia by Morphology 1. -

CD59: a Long-Known Complement Inhibitor Has Advanced to a Blood Group System
B LOOD G ROUP R EVIEW CD59: A long-known complement inhibitor has advanced to a blood group system C. Weinstock, M. Anliker, and I. von Zabern The blood group system number 35 is based on CD59, a 20-kDa by Zalman et al.1 from the group of H.J. Muller-Eberhard in La membrane glycoprotein present on a large number of different Jolla, California, and in 1988 by Schönermark et al. from the cells, including erythrocytes. The major function of CD59 is to group of G.M. Hänsch in Heidelberg (Germany).2 This inhibitor protect cells from complement attack. CD59 binds to complement components C8 and C9 and prevents the polymerization of C9, was published under the designations “HRF (homologous which is required for the formation of the membrane attack restriction factor)” and “C8bp (C8 binding protein),” complex (MAC). Other functions of CD59 in cellular immunity respectively, to describe its properties.1,2 “C8bp” indicates the are less well defined. CD59 is inserted into the membrane by a glycosylphosphatidylinositol (GPI) anchor. A defect of this anchor binding capacity for C8. The name “HRF” points to a species causes lack of this protein from the cell membrane, which leads to incompatibility of this “factor” that provides protection from an enhanced sensitivity towards complement attack. Patients with complement attack more effectively in a homologous (e.g., paroxysmal nocturnal hemoglobinuria (PNH) harbor a varying human erythrocytes as a target of human complement) than in percentage of red blood cell clones with a defect in GPI-anchored proteins, including CD59. The most characteristic symptoms of a heterologous system (e.g., human erythrocytes as a target of this disease are episodes of hemolysis and thromboses. -

SEED Haematology Sysmex Educational Enhancement and Development October 2012
SEED Haematology Sysmex Educational Enhancement and Development October 2012 The red blood cell indices The full blood count has been used in conjunction with the traditional red The complete blood count (CBC) is central to clinical deci- cell indices in order to narrow down the possible causes sion making. This makes it one of the commonest laboratory of anaemia in an individual patient. investigations performed worldwide. Whilst the definition of what constitutes an CBC is influenced by the number Impedance technology and type of parameters measured by different haematology The RBC, HCT and MCV are all closely interrelated as they analysers, the traditional red cell indices that are widely are derived from information obtained from the passage used to classify anaemias are common to all. of cells through the aperture of the impedance channel of an automated haematology analyser. The impedance The laboratory approach to anaemia technology is based on the principle that an electrical field, Anaemia is an extremely common global healthcare prob- created between two electrodes of opposite charge, can lem. However, anaemia is merely a symptom which can be used to count and determine the size of cells. Blood result from a multitude of causes. Effective treatment is cells are poor conductors of electricity. The diluent in which only possible if the underlying cause is correctly identified. they are suspended as they pass through the aperture To this end, several classification systems have been devis- during counting is an isotonic solution which is a good ed. The most useful and widely used classification system conductor of electricity. -

The Widespread Application of Red Cell Survival Sive Red
CLINICAL DETERMINATION OF THE SITES OF RED CELL SEQUESTRATION IN HEMOLYTIC ANEMIAS1 By JAMES H. JANDL, MORTIMER S. GREENBERG, ROBERT H. YONEMOTO, AND WILLIAM B. CASTLE (From the Thorndike Memorial Laboratory and Second and Fourth (Harvard), Medical Services Boston City Hospital, and the Department of Medicine, Harvard Medical School, Boston, Mass.) (Submitted for publication January 30, 1956; accepted April 3, 1956) The widespread application of red cell survival greater than that of other tissues even when cor- techniques has revealed the importance of exces- rection was made for the Cr5l activity of the re- sive red cell destruction in the pathologic physi- sidual red cells. Moreover, the radioactivity of ology of many of the anemias. An increasing ar- the packed red cells removed from the spleen ex- ray of in vitro methods for detecting red cell or ceeded that of a comparable sample of packed red serum abnormalities has provided insight into the cells from the peripheral blood. In order to in- in vivo mechanisms underlying some of these proc- vestigate the possibility that Cr51-labelled red cell esses. In certain disease states the presence of deposition could be determined by measuring body visible or physically measurable alterations of the surface radioactivity, several questions required red cells has permitted detection of the sites and exploration: 1) Are the emanations of Cr5 suit- to some extent of the mechanisms of sequestration able for external body scanning at safe dosage of these cells. Such valuable observations have levels? 2) Does the site of tissue deposition of been made upon pathologic material from patients Cr65 following the intravenous injection of Cr51- with congenital hemolytic anemia (1-5) and labelled red cells necessarily indicate the site of sickle cell anemia (1, 5-7). -

Igm Autoantibody Testing by Latex Agglutination Nephelometry and Elisa in Patients with Rheumatoid Arthritis
European Journal of Molecular & Clinical Medicine ISSN 2515-8260 Volume 07, Issue 08, 2020 Comparative Study Of Rheumatoid Factor - Igm Autoantibody Testing By Latex Agglutination Nephelometry And Elisa In Patients With Rheumatoid Arthritis Dr.C. Devi1, Dr.R. Ravichandran2, Dr. Logeswari Selvaraj3, Dr.S. Ramesh4, Dr.T. Aarthipriya5 1,2,3,4,5Senior Assistant professor Department of Microbiology, Government Kilpauk Medical College, Chennai mail id: [email protected] ABSTRACT Objectives: To test Rheumatoid factor (RF) IgM autoantibody in Patients with Rheumatoid Arthritis by various methods like latex agglutination, Nephelometer and ELISA. Comparative analysis of the sensitivity and specificity of the tests performed. Materials and Methods: The study was conducted for a period of six months from June 2018 to November 2018 in a tertiary care hospital in Chennai. 90 patients attending Rheumatology OPD or admitted in the ward with the diagnosis of Rheumatoid arthritis, satisfying the inclusion and exclusion criteria were taken up for the study. Inclusion criteria: Clinically diagnosed Rheumatoid Arthritis patient as per revised ACR 1987 classification criteria. Duration of symptoms (1yr- early (RA) 1 Yr. (Established RA). Exclusion Criteria: Those with systemic connective tissue diseases like SLE, Scleroderma, MCTD, Sjogren syndrome, those with chronic liver diseases, tuberculosis, subacute Bacterial endocarditis, Pregnancy, Lympho reticular malignancies are excluded for the study. Those with onset 16 years of age are also excluded. Under aseptic precautions about 3ml of blood was collected from each Patient. Rheumatoid factor (RF) IgM was tested for each patient by all three methods Latex agglutination, Nephelometry and ELISA. Results: IgM rheumatoid factor (RF) was detected in the sera of 90 patients with Rheumatoid Arthritis. -

Routine Quantification of Rheumatoid Factor by Rate Nephelometry
Ann Rheum Dis: first published as 10.1136/ard.44.6.379 on 1 June 1985. Downloaded from Annals of the Rheumatic Diseases, 1985, 44, 379-383 Routine quantification of rheumatoid factor by rate nephelometry P J ROBERTS-THOMSON, R McEVOY, T LANGHANS AND J BRADLEY From the Department of Clinical Immunology, Flinders Medical Centre, South Australia 5042 SUMMARY In a cross-sectional study of over 3000 consecutive serum specimens the levels of rheumatoid factor (RF) measured by rate nephelometry (Beckman ICS II) were compared with values obtained by the more traditional methods of sheep cell agglutination (Rose-Waaler) and latex agglutination. Similar values for sensitivity and specificity were found for all three methods for rheumatoid arthritis, with nephelometry giving slightly higher levels of sensitivity for other rheumatic disorders. A significant correlation (r=0-46, p<001) was found between the nephelometric and Rose-Waaler method for 147 consecutive seropositive specimens. Of interest, however, several disparate results were observed, and explanations for these were sought. Longitudinal studies of RF were performed in 49 seropositive patients over a two-year period. The nephelometric method was considered superior compared with the other techniques because of its ability to detect changes in absolute levels at earlier stages and its low interassay coefficient of variance (11%). copyright. We conlcude that the nephelometric technique appears suitable for routine diagnostic use, offers several advantages compared with more traditional methods, and is no more expensive per test specimen than the Rose-Waaler technique. http://ard.bmj.com/ Traditionally, rheumatoid factor (RF) has been interaction of RF with heat aggregated human IgG.