I M M U N O L O G Y Core Notes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lactoferrin and Its Detection Methods: a Review
nutrients Review Lactoferrin and Its Detection Methods: A Review Yingqi Zhang, Chao Lu and Jin Zhang * Department of Chemical and Biochemical Engineering, University of Western Ontario, London, ON N6A 5B9, Canada; [email protected] (Y.Z.); [email protected] (C.L.) * Correspondence: [email protected] Abstract: Lactoferrin (LF) is one of the major functional proteins in maintaining human health due to its antioxidant, antibacterial, antiviral, and anti-inflammatory activities. Abnormal levels of LF in the human body are related to some serious diseases, such as inflammatory bowel disease, Alzheimer’s disease and dry eye disease. Recent studies indicate that LF can be used as a biomarker for diagnosis of these diseases. Many methods have been developed to detect the level of LF. In this review, the biofunctions of LF and its potential to work as a biomarker are introduced. In addition, the current methods of detecting lactoferrin have been presented and discussed. We hope that this review will inspire efforts in the development of new sensing systems for LF detection. Keywords: lactoferrin; biomarkers; immunoassay; instrumental analysis; sensor 1. Introduction Lactoferrin (known as lactotransferrin, LF), with a molecular weight of about 80 kDa, is a functional glycoprotein, which contains about 690 amino acid residues. It was first isolated from bovine milk by Sorensen in 1939 and was first isolated from human milk by Citation: Zhang, Y.; Lu, C.; Zhang, J. Johanson in 1960 [1,2]. The three-dimensional structure of LF has been unveiled by high Lactoferrin and Its Detection resolution X-ray crystallographic analysis, and it consists of two homologous globular lobes Methods: A Review. -
IFM Innate Immunity Infographic
UNDERSTANDING INNATE IMMUNITY INTRODUCTION The immune system is comprised of two arms that work together to protect the body – the innate and adaptive immune systems. INNATE ADAPTIVE γδ T Cell Dendritic B Cell Cell Macrophage Antibodies Natural Killer Lymphocites Neutrophil T Cell CD4+ CD8+ T Cell T Cell TIME 6 hours 12 hours 1 week INNATE IMMUNITY ADAPTIVE IMMUNITY Innate immunity is the body’s first The adaptive, or acquired, immune line of immunological response system is activated when the innate and reacts quickly to anything that immune system is not able to fully should not be present. address a threat, but responses are slow, taking up to a week to fully respond. Pathogen evades the innate Dendritic immune system T Cell Cell Through antigen Pathogen presentation, the dendritic cell informs T cells of the pathogen, which informs Macrophage B cells B Cell B cells create antibodies against the pathogen Macrophages engulf and destroy Antibodies label invading pathogens pathogens for destruction Scientists estimate innate immunity comprises approximately: The adaptive immune system develops of the immune memory of pathogen exposures, so that 80% system B and T cells can respond quickly to eliminate repeat invaders. IMMUNE SYSTEM AND DISEASE If the immune system consistently under-responds or over-responds, serious diseases can result. CANCER INFLAMMATION Innate system is TOO ACTIVE Innate system NOT ACTIVE ENOUGH Cancers grow and spread when tumor Certain diseases trigger the innate cells evade detection by the immune immune system to unnecessarily system. The innate immune system is respond and cause excessive inflammation. responsible for detecting cancer cells and This type of chronic inflammation is signaling to the adaptive immune system associated with autoimmune and for the destruction of the cancer cells. -

C3b Catalog Number: A114 Sizes Available
Name: C3b Catalog Number: A114 Sizes Available: 250 µg/vial Concentration: 1.0 mg/mL (see Certificate of Analysis for actual concentration) Form: Liquid Purity: >90% by SDS-PAGE Buffer: 10 mM sodium phosphate, 145 mM NaCl, pH 7.3 Extinction Coeff. A280 nm = 1.03 at 1.0 mg/mL Molecular Weight: 176,000 Da (2 chains) Preservative: None, 0.22 µm filtered Storage: -70oC or below. Avoid freeze/thaw. Source: Normal human serum (shown by certified tests to be negative for HBsAg and for antibodies to HCV, HIV-1 and HIV-II). Precautions: Use normal precautions for handling human blood products. Origin: Manufactured in the USA. General Description C3b is derived from native C3 upon cleavage and release of C3a. It is prepared by cleavage with the alternative pathway C3 convertase. This is important because only a single bond in C3 is cleaved by the native convertase, while cleavage by other proteases, such as trypsin, results in multiple cleavages at many other sites in the protein. Native human C3b is a glycosylated (~2.8%) polypeptide containing two disulfide-linked chains. C3b is central to the function of all three pathways of complement (Law, S.K.A. and Reid, K.B.M. (1995)). Initiation of each pathway generates proteolytic enzyme complexes (C3 convertases) which are bound to the target surface. These enzymes cleave a peptide bond in C3 releasing the anaphylatoxin C3a and activating C3b. For a brief time (~60 µs) this nascent C3b is capable of reacting with and covalently coupling to hydroxyl groups on the target surface. -

Blocking Properdin Prevents Complement-Mediated Hemolytic Uremic Syndrome and Systemic Thrombophilia
BASIC RESEARCH www.jasn.org Blocking Properdin Prevents Complement-Mediated Hemolytic Uremic Syndrome and Systemic Thrombophilia Yoshiyasu Ueda,1 Takashi Miwa,1 Damodar Gullipalli,1 Sayaka Sato,1 Daisuke Ito,1 Hangsoo Kim,1 Matthew Palmer,2 and Wen-Chao Song1 Departments of 1Systems Pharmacology and Translational Therapeutics and 2Pathology and Laboratory Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania ABSTRACT Background Properdin (P) is a positive regulator of the alternative pathway of complement activation. Although P inhibition is expected and has been shown to ameliorate the alternative pathway of comple- ment-mediated tissue injury in several disease models, it unexpectedly exacerbated renal injury in a murine model of C3 glomerulopathy. The role of P in atypical hemolytic uremic syndrome (aHUS) is uncertain. Methods We blocked P function by genetic deletion or mAb-mediated inhibition in mice carrying a factor H (FH) point mutation, W1206R (FHR/R), that causes aHUS and systemic thrombophilia with high mortality. Results Pdeficiency completely rescued FHR/R mice from premature death and prevented thrombocyto- penia, hemolytic anemia, and renal disease. It also eliminated macrovessel thrombi that were prevalent in FHR/R mice. All mice that received a function-blocking anti-P mAb for 8 weeks survived the experimental period and appeared grossly healthy. Platelet counts and hemoglobin levels were significantly improved in FHR/R mice after 4 weeks of anti-P mAb treatment. One half of the FHR/R mice treated with an isotype control mAb but none of the anti-P mAb-treated mice developed stroke-related neurologic disease. Anti-P mAb-treated FHR/R mice showed largely normal renal histology, and residual liver thrombi were detected in only three of 15 treated mice. -

Levels, B-1 and B-2 Cells, and Antibody Responses Ir Allotype Heterozygous F1 Mice ANN MARIE HAMILTON and JOHN F
Developmental Immunology, 1994, Vol 4, pp. 27-41 (C) 1994 Harwood Academic Publishers GmbH Reprints available directly from the publisher Printed in Singapore Photocopying permitted by license only Effects of IgM Allotype Suppression on Serum IgM Levels, B-1 and B-2 Cells, and Antibody Responses ir Allotype Heterozygous F1 Mice ANN MARIE HAMILTON and JOHN F. KEARNEY" Division of Developmental and Clinical Immunology, Department of Microbiology, University of Alabama, Birmingham, Alabama 35294 IgM allotype heterozygous F1 mice were independently suppressed for Igh6a or Igh6b to evaluate the contribution of B-1 and B-2 cells to natural serum IgM levels and Ab responses. B-2 B cells expressing IgM of the suppressed allotype were evident in the spleens of suppressed mice 4 to 6 weeks after cessation of the suppression regimen, whereas B-1 B cells of the suppressed allotype were undetectable for up to 9 months. Although serum IgM of the suppressed allotype was initially depleted in mice suppressed for either allotype, by 7 months of age, there were detectable levels of IgM of the suppressed allotype in the serum; however, the levels were significantly below that found in nonsuppressed mice. When mice were immunized with either the T-independent or T-dependent form of phosphorylcholine, those suppressed for either allotype, and consequently depleted of B-1 B cells of that allotype, did not respond with phosphorylcholine-specific IgM of the suppressed allotype. In contrast, when mice were immunized with 1-3 dextran, the Igh6a allotype-suppressed mice were able to produce dextran-specific IgM of that allotype. -

Role of Igg3 in Infectious Diseases
Review Role of IgG3 in Infectious Diseases 1 2 1 1, Timon Damelang, Stephen J. Rogerson, Stephen J. Kent, and Amy W. Chung * IgG3 comprises only a minor fraction of IgG and has remained relatively under- Highlights studied until recent years. Key physiochemical characteristics of IgG3 include IgG3 has been associated with enhanced control or protection against an elongated hinge region, greater molecular flexibility, extensive polymor- a range of intracellular bacteria, para- phisms, and additional glycosylation sites not present on other IgG subclasses. sites, and viruses. These characteristics make IgG3 a uniquely potent immunoglobulin, with the IgG3 Abs are potent mediators of potential for triggering effector functions including complement activation, effector functions, including enhanced antibody (Ab)-mediated phagocytosis, or Ab-mediated cellular cytotoxicity ADCC, opsonophagocytosis, comple- ment activation, and neutralization, (ADCC). Recent studies underscore the importance of IgG3 effector functions compared with other IgG subclasses. against a range of pathogens and have provided approaches to overcome IgG3-associated limitations, such as allotype-dependent short Ab half-life, and Future Ab-based therapeutics and vaccines should consider utilizing excessive proinflammatory activation. Understanding the molecular and func- IgG3, based on features of enhanced tional properties of IgG3 may facilitate the development of improved Ab-based functional capacity. immunotherapies and vaccines against infectious diseases. Investigating the impact of glycosyla- tion patterns and allotypes on IgG3 Human IgG3 an Understudied but Highly Potent Immunoglobulin function may expand our understand- Antibodies (Abs) play a major role in protection against infections by binding to and inactivating ing of IgG3 responses and their ther- apeutic potential. invading pathogens. -

Innate Immunity in the Lung: How Epithelial Cells Fight Against
Copyright #ERSJournals Ltd 2004 EurRespir J 2004;23: 327– 333 EuropeanRespiratory Journal DOI: 10.1183/09031936.03.00098803 ISSN0903-1936 Printedin UK –allrights reserved REVIEW Innateimmunity in the lung: how epithelialcells ght against respiratorypathogens R. Bals*,P.S. Hiemstra # Innateimmunity in the lung: how epithelial cells ® ghtagainst respiratory pathogens *Deptof Internal Medicine, Division of R Bals, P S Hiemstra #ERS JournalsLtd 2004 PulmonaryMedicine, Hospital of the Uni- versityof Marburg, Philipps-University, ABSTRACT: Thehuman lung is exposed to a largenumber of airborne pathogens as a # resultof the daily inhalation of 10,000 litres of air Theobservation that respiratory Marburg,Germany; Deptof Pulmonology, LeidenUniversity Medical Center, Leiden, infectionsare nevertheless rare is testimony to the presence of anef®cient host defence TheNetherlands systemat the mucosal surface of the lung Theairway epithelium is strategically positioned at the interface with the Correspondence:P S Hiemstra,Dept of environment,and thus plays a keyrole in this host defence system Recognition Pulmonology,Leiden University Medical systemsemployed by airwayepithelial cells to respond to microbialexposure include the Center, P O Box9600, 2300 RC Leiden, The actionof the toll-like receptors Netherlands Theairway epithelium responds to such exposure by increasing its production of Fax:31 715266927 mediatorssuch as cytokines, chemokines and antimicrobial peptides Recent® ndings E-mail: p s hiemstra@lumc nl indicatethe importance of these peptides -

Practice Guideline: Disease-Modifying Therapies for Adults with Multiple Sclerosis
Practice guideline: Disease-modifying therapies for adults with multiple sclerosis Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology Alexander Rae-Grant, MD1; Gregory S. Day, MD, MSc2; Ruth Ann Marrie, MD, PhD3; Alejandro Rabinstein, MD4; Bruce A.C. Cree, MD, PhD, MAS5; Gary S. Gronseth, MD6; Michael Haboubi, DO7; June Halper, MSN, APN-C, MSCN8; Jonathan P. Hosey, MD9; David E. Jones, MD10; Robert Lisak, MD11; Daniel Pelletier, MD12; Sonja Potrebic, MD, PhD13; Cynthia Sitcov14; Rick Sommers, LMSW15; Julie Stachowiak, PhD16; Thomas S.D. Getchius17; Shannon A. Merillat, MLIS18; Tamara Pringsheim, MD, MSc19 1. Department of Neurology, Cleveland Clinic, OH 2. Department of Neurology, Charles F. and Joanne Knight Alzheimer Disease Research Center, Washington University in St. Louis, MO 3. Departments of Medicine and Community Health Sciences, Max Rady College of Medicine, Rady Faculty of Health Sciences, University of Manitoba, Winnipeg, Canada 4. Department of Neurology, Mayo Clinic, Rochester, MN 5. UCSF Weill Institute for Neurosciences, Department of Neurology, University of California, San Francisco 6. Department of Neurology, Kansas University Medical Center, Kansas City 7. Department of Neurology, School of Medicine, University of Louisville, KY 8. Consortium of Multiple Sclerosis Centers, Hackensack, NJ 9. Department of Neuroscience, St. Luke’s University Health Network, Bethlehem, PA 10. Department of Neurology, School of Medicine, University of Virginia, Charlottesville 11. Consortium of Multiple Sclerosis Centers, Hackensack, NJ, and Department of Neurology, School of Medicine, Wayne State University, Detroit, MI 12. Department of Neurology, Keck School of Medicine, University of Southern California, Los Angeles 13. Neurology Department, Southern California Permanente Medical Group, Kaiser, Los Angeles 14. -

Monoclonal Antibody Playbook
Federal Response to COVID-19: Monoclonal Antibody Clinical Implementation Guide Outpatient administration guide for healthcare providers 2 SEPTEMBER 2021 1 Introduction to COVID-19 Monoclonal Antibody Therapy 2 Overview of Emergency Use Authorizations 3 Site and Patient Logistics Site preparation Patient pathways to monoclonal administration 4 Team Roles and Responsibilities Leadership Administrative Clinical Table of 5 Monoclonal Antibody Indications and Administration Indications Contents Preparation Administration Response to adverse events 6 Supplies and Resources Infrastructure Administrative Patient Intake Administration 7 Examples: Sites of Administration and Staffing Patterns 8 Additional Resources 1 1. Introduction to Monoclonal Therapy 2 As of 08/13/21 Summary of COVID-19 Therapeutics 1 • No Illness . Health, no infections • Exposed Asymptomatic Infected . Scope of this Implementation Guide . Not hospitalized, no limitations . Monoclonal Antibodies for post-exposure prophylaxis (Casirivimab + Imdevimab (RGN)) – EUA Issued. • Early Symptomatic . Scope of this Implementation Guide . Not hospitalized, with limitations . Monoclonal Antibodies for treatment (EUA issued): Bamlanivimab + Etesevimab1 (Lilly) Casirivimab + Imdevimab (RGN) Sotrovimab (GSK/Vir) • Hospital Adminission. Treated with Remdesivir (FDA Approved) or Tocilizumab (EUA Issued) . Hospitalized, no acute medical problems . Hospitalized, not on oxygen . Hospitlaized, on oxygen • ICU Admission . Hospitalized, high flow oxygen, non-invasive ventilation -
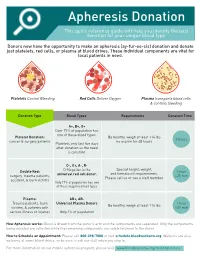
Apheresis Donation This Quick Reference Guide Will Help You Identify the Best Donation for Your Unique Blood Type
Apheresis Donation This quick reference guide will help you identify the best donation for your unique blood type. Donors now have the opportunity to make an apheresis (ay-fur-ee-sis) donation and donate just platelets, red cells, or plasma at blood drives. These individual components are vital for local patients in need. Platelets Control Bleeding Red Cells Deliver Oxygen Plasma transports blood cells & controls bleeding Donation Type Blood Types Requirements Donation Time A+, B+, O+ Over 75% of population has one of these blood types. Platelet Donation: Be healthy, weigh at least 114 lbs 2 hours cancer & surgery patients no aspirin for 48 hours Platelets only last five days after donation so the need is constant. O-, O+, A-, B- Special height, weight, Double Red: O-Negative is the 1 hour and hematocrit requirements. surgery, trauma patients, universal red cell donor. +25 min Please call us or see a staff member accident, & burn victims Only 17% of population has one of these negative blood types Plasma: AB+, AB- Trauma patients, burn Universal Plasma Donors 1 hour Be healthy, weigh at least 114 lbs victims, & patients with +30 min serious illness or injuries Only 4% of population How Apheresis works: Blood is drawn from the donor’s arm and the components are separated. Only the components being donated are collected while the remaining components are safely returned to the donor How to Schedule an Appointment: Please call 800-398-7888 or visit schedule.bloodworksnw.org. Walk-ins are also welcome at some blood drives, so be sure to ask our staff when you stop in. -

Serotyping of Bdellovibrios by Agglutination and Indirect Immunofluorescence
INTERNATIONAL JOURNAL OF SYSTEMATICBACT~RIOLOGY. Oct. 1983, p. 816-821 Vol. 33, No. 4 0020-7713/83/040816-06$02.00/0 Copyright 0 1983, Internationnl Union of Microbiologicdl Societie\ Serotyping of Bdellovibrios by Agglutination and Indirect Immunofluorescence M. E. SCHELLING’* AND S. F. CONTI’ Department Biology, Texus A &M Unitwsit?, College Station, Texus 77843‘ und University of Mussa ch us et t s , A rn ii erst, Muss N c h iiset t J 0 1 002’ A comparative serological study was undertaken to clarify the uncertain interrelationships among various bdellovibrios. A total of 17 predacious (host- dependent) strains and 12 nonpredacious strains were serotyped. Strains were grouped into serotypes on the basis of positive agglutination and strongly positive fluorescence cross-reactions. The results of division of the strains into serogroups by these two methods were identical. Bdellovihrio starrii strain A3.12 and Bdellovibrio stolpii strain UKi2 were found to be antigenically distinct from each other and from the group of strains comprising Bdellovihrio hacteriovorus; the latter was found to be an antigenically heterogeneous group consisting of at least nine serogroups. All nonpredacious strains were found to be related antigenically to their obligately predacious counterparts. An antigen(s) common to all of the Bdellovibrio strains examined was exhibited as weakly positive fluorescence. Selected strains of Bdellovihrio were studied by Ouchterlony immunodiffusion, which confirmed the serogrouping results and provided a technically simple method of serotyping. Further immunochemical characterization of the strains of Bdellovihrio in conjunction with increased knowledge of other differences among strains will likely result in the formation of additional species of Bdellovihrio. -

B Cell Checkpoints in Autoimmune Rheumatic Diseases
REVIEWS B cell checkpoints in autoimmune rheumatic diseases Samuel J. S. Rubin1,2,3, Michelle S. Bloom1,2,3 and William H. Robinson1,2,3* Abstract | B cells have important functions in the pathogenesis of autoimmune diseases, including autoimmune rheumatic diseases. In addition to producing autoantibodies, B cells contribute to autoimmunity by serving as professional antigen- presenting cells (APCs), producing cytokines, and through additional mechanisms. B cell activation and effector functions are regulated by immune checkpoints, including both activating and inhibitory checkpoint receptors that contribute to the regulation of B cell tolerance, activation, antigen presentation, T cell help, class switching, antibody production and cytokine production. The various activating checkpoint receptors include B cell activating receptors that engage with cognate receptors on T cells or other cells, as well as Toll-like receptors that can provide dual stimulation to B cells via co- engagement with the B cell receptor. Furthermore, various inhibitory checkpoint receptors, including B cell inhibitory receptors, have important functions in regulating B cell development, activation and effector functions. Therapeutically targeting B cell checkpoints represents a promising strategy for the treatment of a variety of autoimmune rheumatic diseases. Antibody- dependent B cells are multifunctional lymphocytes that contribute that serve as precursors to and thereby give rise to acti- cell- mediated cytotoxicity to the pathogenesis of autoimmune diseases