Cell and Tissue Structure in Animals and Plants - M.K
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chapter 5: the Shoot System I: the Stem
Chapter 5 The Shoot System I: The Stem THE FUNCTIONS AND ORGANIZATION OF THE SHOOT SYSTEM PRIMARY GROWTH AND STEM ANATOMY Primary Tissues of Dicot Stems Develop from the Primary Meristems The Distribution of the Primary Vascular Bundles Depends on the Position of Leaves Primary Growth Differs in Monocot and Dicot Stems SECONDARY GROWTH AND THE ANATOMY OF WOOD Secondary Xylem and Phloem Develop from Vascular Cambium Wood Is Composed of Secondary Xylem Gymnosperm Wood Differs from Angiosperm Wood Bark Is Composed of Secondary Phloem and Periderm Buds Are Compressed Branches Waiting to Elongate Some Monocot Stems Have Secondary Growth STEM MODIFICATIONS FOR SPECIAL FUNCTIONS THE ECONOMIC VALUE OF WOODY STEMS SUMMARY ECONOMIC BOTANY: How Do You Make A Barrel? 1 KEY CONCEPTS 1. The shoot system is composed of the stem and its lateral appendages: leaves, buds, and flowers. Leaves are arranged in different patterns (phyllotaxis): alternate, opposite, whorled, and spiral. 2. Stems provide support to the leaves, buds, and flowers. They conduct water and nutrients and produce new cells in meristems (shoot apical meristem, primary and secondary meristems). 3. Dicot stems and monocot stems are usually different. Dicot stems tend to have vascular bundles distributed in a ring, whereas in monocot stems they tend to be scattered. 4. Stems are composed of the following: epidermis, cortex and pith, xylem and phloem, and periderm. 5. Secondary xylem is formed by the division of cells in the vascular cambium and is called wood. The bark is composed of all of the tissues outside the vascular cambium, including the periderm (formed from cork cambium) and the secondary phloem. -
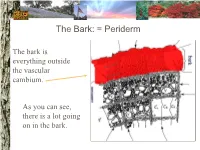
The Bark: = Periderm
The Bark: = Periderm The bark is everything outside the vascular cambium. As you can see, there is a lot going on in the bark. The Bark: periderm: phellogen (cork cambium): The phellogen is the region of cell division that forms the periderm tissues. Phellogen development influences bark appearance. The Bark: periderm: phellem (cork): Phellem replaces the epidermis as the tree increases in girth. Photosynthesis can take place in some trees both through the phellem and in fissures. The Bark: periderm: phelloderm: Phelloderm is active parenchyma tissue. Parenchyma cells can be used for storage, photosynthesis, defense, and even cell division! The Bark: phloem: Phloem tissue makes up the inner bark. However, it is vascular tissue formed from the vascular cambium. The Bark: phloem: sieve tube elements: Sieve tube elements actively transport photosynthates down the stem. Conifers have sieve cells instead. The cambium: The cambium is the primary meristem producing radial growth. It forms the phloem & xylem. The Xylem (wood): The xylem includes everything inside the vascular cambium. The Xylem: a growth increment (ring): The rings seen in many trees represent one growth increment. Growth rings provide the texture seen in wood. The Xylem: vessel elements: Hardwood species have vessel elements in addition to trachieds. Notice their location in the growth rings of this tree The Xylem: fibers: Fibers are cells with heavily lignified walls making them stiff. Many fibers in sapwood are alive at maturity and can be used for storage. The Xylem: axial parenchyma: Axial parenchyma is living tissue! Remember that parenchyma cells can be used for storage and cell division. -

Tree Anatomy Stems and Branches
Tree Anatomy Series WSFNR14-13 Nov. 2014 COMPONENTSCOMPONENTS OFOF PERIDERMPERIDERM by Dr. Kim D. Coder, Professor of Tree Biology & Health Care Warnell School of Forestry & Natural Resources, University of Georgia Around tree roots, stems and branches is a complex tissue. This exterior tissue is the environmental face of a tree open to all sorts of site vulgarities. This most exterior of tissue provides trees with a measure of protection from a dry, oxidative, heat and cold extreme, sunlight drenched, injury ridden site. The exterior of a tree is both an ecological super highway and battle ground – comfort and terror. This exterior is unique in its attributes, development, and regeneration. Generically, this tissue surrounding a tree stem, branch and root is loosely called bark. The tissues of a tree, outside or more exterior to the xylem-containing core, are varied and complexly interwoven in a relatively small space. People tend to see and appreciate the volume and physical structure of tree wood and dismiss the remainder of stem, branch and root. In reality, tree life is focused within these more exterior thin tissue sets. Outside of the cambium are tissues which include transport cells, structural support cells, generation cells, and cells positioned to help, protect, and sustain other cells. All of this life is smeared over the circumference of a predominately dead physical structure. Outer Skin Periderm (jargon and antiquated term = bark) is the most external of tree tissues providing protection, water conservation, insulation, and environmental sensing. Periderm is a protective tissue generated over and beyond live conducting and non-conducting cells of the food transport system (phloem). -

Eudicots Monocots Stems Embryos Roots Leaf Venation Pollen Flowers
Monocots Eudicots Embryos One cotyledon Two cotyledons Leaf venation Veins Veins usually parallel usually netlike Stems Vascular tissue Vascular tissue scattered usually arranged in ring Roots Root system usually Taproot (main root) fibrous (no main root) usually present Pollen Pollen grain with Pollen grain with one opening three openings Flowers Floral organs usually Floral organs usually in in multiples of three multiples of four or five © 2014 Pearson Education, Inc. 1 Reproductive shoot (flower) Apical bud Node Internode Apical bud Shoot Vegetative shoot system Blade Leaf Petiole Axillary bud Stem Taproot Lateral Root (branch) system roots © 2014 Pearson Education, Inc. 2 © 2014 Pearson Education, Inc. 3 Storage roots Pneumatophores “Strangling” aerial roots © 2014 Pearson Education, Inc. 4 Stolon Rhizome Root Rhizomes Stolons Tubers © 2014 Pearson Education, Inc. 5 Spines Tendrils Storage leaves Stem Reproductive leaves Storage leaves © 2014 Pearson Education, Inc. 6 Dermal tissue Ground tissue Vascular tissue © 2014 Pearson Education, Inc. 7 Parenchyma cells with chloroplasts (in Elodea leaf) 60 µm (LM) © 2014 Pearson Education, Inc. 8 Collenchyma cells (in Helianthus stem) (LM) 5 µm © 2014 Pearson Education, Inc. 9 5 µm Sclereid cells (in pear) (LM) 25 µm Cell wall Fiber cells (cross section from ash tree) (LM) © 2014 Pearson Education, Inc. 10 Vessel Tracheids 100 µm Pits Tracheids and vessels (colorized SEM) Perforation plate Vessel element Vessel elements, with perforated end walls Tracheids © 2014 Pearson Education, Inc. 11 Sieve-tube elements: 3 µm longitudinal view (LM) Sieve plate Sieve-tube element (left) and companion cell: Companion cross section (TEM) cells Sieve-tube elements Plasmodesma Sieve plate 30 µm Nucleus of companion cell 15 µm Sieve-tube elements: longitudinal view Sieve plate with pores (LM) © 2014 Pearson Education, Inc. -

+ Complex Tissues
+ Complex Tissues ! Complex tissues are made up of two or more cell types. ! Xylem - Chief conducting tissue for water and minerals absorbed by the roots. ! Vessels - Made of vessel elements. ! Long tubes open at each end. ! Tracheids - Tapered at the ends with pits that allow water passage between cells. ! Rays - Lateral conduction. + Complex Tissues - Xylem ! Tracheids ! Long, thin cells with pointed ends that conduct water vertically ! Line up in columns like pipes by overlapping their tapered ends ! die when reach maturity ! Water conducted through tubes made up of tracheid cell walls ! Wherever two ends join, small holes in the cell wall called pits line up to allow water to flow from one tracheid to another. + Complex Tissues - Xylem ! Pits always occur in pairs so that a pair of pits lines up on either side of the middle lamella, or center layer, of the cell wall. + Complex Tissues - Xylem !Vessel elements !barrel-shaped cells with open ends that conduct water vertically. !line up end to end forming columns, called vessels, that conduct water. !Some have completely open ends, while others have narrow strips of cell wall material that partially covers the ends !Die at maturity, like tracheids. + Complex Tissues - Xylem ! Ray cells………. ! Long lived parenchyma cells that extend laterally like the spokes of a wheel from the center of a woody stem out towards the exterior of the stem ! alive at maturity ! Transport materials horizontally from center outward + Complex Tissues - Xylem ! Xylem fibers – ! long, thin sclerenchyma cells ! Xylem parenchyma cells- that run parallel to the ! Living cells vessel element ! Distributed among tracheids and vessels ! Help strengthen and support xylem ! Store water and nutrients + Complex Tissues - Phloem ! Phloem brings sugar [glucose from photosynthesis] from the leaves to all parts of the plant body. -

Tree Anatomy Stems and Branches
Tree Anatomy: STEM COMPONENTS Dr. Kim D. Coder, Professor of Tree Biology & Health Care, Warnell School, UGA Trees attempt to occupy more space and control available resources through cell divisions and cell expansion. From the end of a shoot tip to the end of a root tip, trees elongate. Along this axis of growth, trees also expand radially, which is termed “secondary growth.” Tree growth is initiated in the shoot tips (growing points / buds), root tips, vascular cambium (i.e. or simply cambium), and phellogen which generates periderm. The first two meristems are primary generation systems and the second two are termed secondary meristems. Cambial activity and xylem formation, as seen as visible increases in growth increment volume are radial growth, and reviewed here. Cross-Section Tree stem anatomy is usually visualized in cross-section. In this form, two-demensional “layers” or “rings,” and tissue types can be identified. More difficult is to visualize these two-dimensional cross- sections and incorporate them into a three dimensional object which changes and grows over time. Moving from outside to inside a stem, tissues encountered include those associated with periderm, secondary cortex, phloem, xylem, and pith. A traditional means of describing a mature tree stem cross-section defines which tissue type is cut first, second and third using a handsaw. A list of generic tissues in stems from outside to inside could include: Figure 1. epidermis and primary cortex = exterior primary protective tissues quickly rent, torn, and discarded with secondary growth (expansion of girth by lateral meristems – vascular cambium and phellogen). periderm = a multiple layer tissue responsible for tree protection and water conservation generated over the exterior of a tree from shoot to root, and produced by the phellogen (cork cambium – lateral meristem). -

Petrified Wood: the Anatomy of Arborescent Plant Life Through Time
The Anatomy of Arborescent Plant Life Through Time Mike Viney Collectors of petrified wood focus on permineralized plant material related to arborescent (tree-like) plant life. Evidence for the first fossil forest occurs in the Devonian. Fossil forest composition changes through geologic time, reflecting variety in evolutionary strategies for constructing a tree form. It is helpful and informative to study the anatomy of various trunk designs. Evolutionary adaptations for trunk structure can be recognized by the arrangement of tissues and organs. A quick survey of plant organs and tissues will enhance our discussion of the various evolutionary strategies for constructing a tree form. Plants are made of four types of organs: roots, stems, leaves, and reproductive structures. In turn, these organs are composed of three basic tissue systems: the ground tissue system, the vascular tissue system, and the dermal tissue system. Ground tissues including parenchyma, collenchyma and sclerenchyma are involved in photosynthesis, storage, secretion, transport, and structure. Parenchyma tissue produces all other tissues. Living parenchyma cells are involved in photosynthesis, storage, secretion, regeneration and in the movement of water and food. Parenchyma cells are typically spherical to cube shaped. Collenchyma tissue provides structural support for young growing organs. Living collenchyma cells are elongated cylinders and help to make up the familiar string-like material in celery stalks and leaf petioles. Sclerenchyma tissue provides support for primary and secondary plant bodies. Sclerenchyma cells often have lignified secondary walls and lack protoplasm at maturity. Elongated slender sclerenchyma cells known as fibers make up well known fibrous material such as hemp, jute, and flax. -

Cork Cambium - Produces Cork (Outer Most Layer of Bark) Pine Tree W/ 8 Cotyledons!
Plantae Seed Plants Vascular Plants • Formation of vascular tissue – Xylem (water) – Phloem (food) – True leaves, roots, and stems • Lignin • ____________ generation dominate Alternation of Generation Alternation of Generation • Sporophyte dependent on gametophyte – mosses • Large sporophyte and small independent gametophyte – ferns • Gametophyte dependent on sporophyte – seed plants Why be Sporophyte Dominant? • Reduced mutations – UV light harmful to DNA – Diploid (2n) form copes better with mutations • two alleles Why Retain Gametophyte Generation? • Ability to screen alleles – doesn’t require a large amount of energy • Sporophyte embryos rely on some gametophyte tissue Seeds • A seed is a sporophyte in a package – spores are only single cells – packaged with food • All seed plants are _____________ (more than one kind of spore) – megasporangia – microsporangia From Ovule to Seed Develops from megaspore Whole structure Embryo, food supply, protective coat Overview of Seed Plants • Produce Seeds – Can remain dormant for years – Pollination replaces swimming sperm • Gametophyte generation reduced – Gymnosperms lack antheridium – Angiosperms lack both archegonium and antheridium Phylogeny Gymnosperms (Naked Seed) • Division: Cycadophyta • Division: Ginkgophyta • Division: Gnetophyta • Division: Coniferophyta Ginkgophyta • Ginkgo or Maidenhair Tree • Characteristic leaves • Only one species • Only ______ are planted Cycadophyta • Cycads • Palm-like plants – Sago Palms • Leaves in cluster at top of trunks • True __________ Gnetophyta • 3 -

CHAPTER 3 COMPARATIVE ANATOMY of BARK on BRANCHES and RAMETS of COMMON DICOTYLEDONOUS TREES and WOODY SHRUBS. 3.1. Introduction
1 CHAPTER 3 COMPARATIVE ANATOMY OF BARK ON BRANCHES AND RAMETS OF COMMON DICOTYLEDONOUS TREES AND WOODY SHRUBS. 3.1. Introduction 3.1.1 The goal This is an introduction into the interpretation of the anatomy of bark of some common dicot branches and woody shrubs ramets that may have been used in ethnographic or archaeological artifacts. Branches with their bark still on have been used for many ethnographic purposes, mainly with basket making, As well as mats on floors and bundles in festive dances. Bark alone has been was used for basket decoration, such as imbrications. Many components of bark species have been used for dyes and medicinal purposes. In archaeological and ethnological studies there may be a need to identify species origin of bark fragments as well as intact bark, thus the purpose of the following anatomical study of the bark on branches.. The purpose of the following is to give a working vocabulary of the important anatomical features of the bark. The information presented allows identification of some common branch species or genera of American North West Coast. In many cases the species studied have common characteristic of other species of the genera. The identification of species in a genera, in many cases, depends on morphological features such as flower or leaf shape, color, etc. There is nothing in the literature that gives a comparative analysis of this group of species for identification, but there is some literature on the individual species. The approach is first to obtain the visual knowledge of the main features of barks. -

Plant Structure and Growth
Plant Structure And Growth The Plant Body is Composed of Cells and Tissues • Tissue systems (Like Organs) – made up of tissues • Made up of cells Plant Tissue Systems • ____________________Ground Tissue System Ø photosynthesis Ø storage Ø support • ____________________Vascular Tissue System Ø conduction Ø support • ___________________Dermal Tissue System Ø Covering Ground Tissue System • ___________Parenchyma Tissue • Collenchyma Tissue • Sclerenchyma Tissue Parenchyma Tissue • Made up of Parenchyma Cells • __________Living Cells • Primary Walls • Functions – photosynthesis – storage Collenchyma Tissue • Made up of Collenchyma Cells • Living Cells • Primary Walls are thickened • Function – _Support_____ Sclerenchyma Tissue • Made up of Sclerenchyma Cells • Usually Dead • Primary Walls and secondary walls that are thickened (lignin) • _________Fibers or _________Sclerids • Function – Support Vascular Tissue System • Xylem – H2O – ___________Tracheids – Vessel Elements • Phloem - Food – Sieve-tube Members – __________Companion Cells Xylem • Tracheids – dead at maturity – pits - water moves through pits from cell to cell • Vessel Elements – dead at maturity – perforations - water moves directly from cell to cell Phloem Sieve-tube member • Sieve-tube_____________ members – alive at maturity – lack nucleus – Sieve plates - on end to transport food • _____________Companion Cells – alive at maturity – helps control Companion Cell (on sieve-tube the side) member cell Dermal Tissue System • Epidermis – Single layer, tightly packed cells – Complex -

Anatomy of Flowering Plants
Anatomy of Flowering Plants Anatomy definition: is the branch of biology concerned with the study of the structure of organisms and their parts. Difference between Morphology and Anatomy: o Plant morphology or phytomorphology is the study of the physical form and external structure of plants, whereas plant anatomy is the study of the internal plant structure, mostly at the cellular/microscopic level. Cell – is the smallest structural and functional unit of an organism, which is typically microscopic and consist of cytoplasm and nucleus enclosed in a membrane. Difference between Protoplasm and cytoplasm: o Protoplasm is the colourless material consisting of the living part of a cell, including the cytoplasm, nucleus, and other organelles. Cytoplasm is the fluid that consist of all of the content outside the nucleus and enclosed within the cell membrane of a cell. The tissue A group of cells having a common origin and usually performing common function are called tissues. Plant Tissue Meristematic Tissue Permanent Tissue 1. Apical meristem Simple permanent tissue Complex permanent tissue 2. Intercalary 1. Parenchyma 1. Xylem 3. Lateral 2. Collenchyma 2. Phloem 3. Sclerenchyma Meristematic Tissues or Meristems (1) They contain immature and young cells and are capable of repeated divisions. (2) Intercellular spaces are not present in meristematic tissue. (3) They contain a homogeneous thin wall. (4) They contain large nuclei associated with abundant cytoplasm. (5) They are metabolically very active but they do not store food material. (6) Only proto-plastids are present instead of plastids, chloroplast absent. (7) Dense cytoplasm is present which contains several premature mitochondria. (8) Vacuoles are absent. -

Botany of Trees 2
Botany of Trees California’s Part 2: How Wood Forms Iconic Flora North Coast California Na.ve Plant Society Free Webinar 7 PM Tomorrow Night Dr. Ma9 Ri9er h9ps://us04web.zoom.us/webinar/register/ WN_s4i_BmnlTryUe_7t45a9Qw Botany of Trees Tuesday, April 14th: IntroducTon to trees, growth, development, leaves, and morphology • Five part series -Tuesdays, 11 AM to 12:30 PM • Each will be ~1 hour with 30 minutes of ques.ons Tuesday, April 28th: Tree names, Tuesday, April 21st: Trunks, branches, diversity, and why names change tree form, branching pa9erns, and shape, and how wood forms Tuesday, May 5th: Water in trees, Tuesday, May 12th: ReproducTon, photosynthesis, and respiraTon flower formaTon, fruit, and seeds All aboveground plant structures are: Interpret the sharp structure: stems, leaves, or buds Modified leaf - spine bud becoming a new branch Pointy Structures on Plants Big Leaf Maple (Acer macrophyllum) Thorn Spine Prickle modified branch modified leaf epidermal that comes from that comes from outgrowths that an axillary bud below the occur at random on axillary bud the stem (not necessarily at nodes) Big Leaf Maple (Acer Pine Leaves macrophyllum) • Leaves of two kinds: primary scales and secondary needles • Primary leaves of pines are membranous scales • A set number of needle leaves are produced on short branches (fascicles) • Each bundle (fascicle) of needles is surrounded by membranous bud scales Leaf Leaf Leaf Stump SprouTng & Epicormic Growth Meristems: where growth occurs • Apical Meristems • Form primary .ssues • Increase in