Anatomy of Flowering Plants
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bark Anatomy and Cell Size Variation in Quercus Faginea
Turkish Journal of Botany Turk J Bot (2013) 37: 561-570 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1201-54 Bark anatomy and cell size variation in Quercus faginea 1,2, 2 2 2 Teresa QUILHÓ *, Vicelina SOUSA , Fatima TAVARES , Helena PEREIRA 1 Centre of Forests and Forest Products, Tropical Research Institute, Tapada da Ajuda, 1347-017 Lisbon, Portugal 2 Centre of Forestry Research, School of Agronomy, Technical University of Lisbon, Tapada da Ajuda, 1349-017 Lisbon, Portugal Received: 30.01.2012 Accepted: 27.09.2012 Published Online: 15.05.2013 Printed: 30.05.2013 Abstract: The bark structure of Quercus faginea Lam. in trees 30–60 years old grown in Portugal is described. The rhytidome consists of 3–5 sequential periderms alternating with secondary phloem. The phellem is composed of 2–5 layers of cells with thin suberised walls and narrow (1–3 seriate) tangential band of lignified thick-walled cells. The phelloderm is thin (2–3 seriate). Secondary phloem is formed by a few tangential bands of fibres alternating with bands of sieve elements and axial parenchyma. Formation of conspicuous sclereids and the dilatation growth (proliferation and enlargement of parenchyma cells) affect the bark structure. Fused phloem rays give rise to broad rays. Crystals and druses were mostly seen in dilated axial parenchyma cells. Bark thickness, sieve tube element length, and secondary phloem fibre wall thickness decreased with tree height. The sieve tube width did not follow any regular trend. In general, the fibre length had a small increase toward breast height, followed by a decrease towards the top. -

Differentiation of Epidermis with Reference to Stomata
Unit : 3 Differentiation of Epidermis with Reference to Stomata LESSON STRUCTURE 3.0 Objective 3.1 Introduction 3.2 Epidermis : Basic concept, its function, origin and structure 3.3 Stomatal distribution, development and classification. 3.4 Questions for Exercise 3.5 Suggested Readings 3.0 OBJECTIVE The epidermis, being superficial or outermost layer of cells, covers the entire plant body. It includes structures like stomata and trichomes. Distribution of stomata in epidermis depends on Ontogeny, number of subsidiary cells, separation of guard cells and different taxonomic ranks (classes, families and species). 3.1 INTRODUCTION The internal organs of plants are covered by a well developed tissue system, the epidermal or integumentary system. The epidermis modifies itself to cope up with natural surroundings, since, it is in direct contact with the environment. It protects the inner tissues from any adverse natural calamities like high temperature, desiccation, mechanical injury, excessive illumination, external infection etc. In some plants epidermis may persist throughout the life, while in others it is replaced by periderm. Although the epiderm usually arises from the outer most tunica layer, which thus coincides with Hanstein’s dermatogen, the underlying tissues may have their origin in the tunica or the corpus or both, depending on plant species and the number of tunica layers, explained by Schmidt (1924). ( 38 ) Differentiation of Epidermis with Reference to Stomata 3.2 EPIDERMIS Basic Concept The term epidermis designates the outer most layer of cells on the primary plant body. The word is derived from two Greek words ‘epi’ means upon and ‘derma’ means skin. Through the history of development of plant morphology the concept of the epidermis has undergone changes, and there is still no complete uniformity in the application of the term. -

Stoma and Peristomal Skin Care: a Clinical Review Early Intervention in Managing Complications Is Key
WOUND WISE 1.5 HOURS CE Continuing Education A series on wound care in collaboration with the World Council of Enterostomal Therapists Stoma and Peristomal Skin Care: A Clinical Review Early intervention in managing complications is key. ABSTRACT: Nursing students who don’t specialize in ostomy care typically gain limited experience in the care of patients with fecal or urinary stomas. This lack of experience often leads to a lack of confidence when nurses care for these patients. Also, stoma care resources are not always readily available to the nurse, and not all hospitals employ nurses who specialize in wound, ostomy, and continence (WOC) nursing. Those that do employ WOC nurses usually don’t schedule them 24 hours a day, seven days a week. The aim of this article is to provide information about stomas and their complications to nurses who are not ostomy specialists. This article covers the appearance of a normal stoma, early postoperative stoma complications, and later complications of the stoma and peristomal skin. Keywords: complications, ostomy, peristomal skin, stoma n 46 years of clinical practice, I’ve encountered article covers essential information about stomas, many nurses who reported having little educa- stoma complications, and peristomal skin problems. Ition and even less clinical experience with pa- It is intended to be a brief overview; it doesn’t provide tients who have fecal or urinary stomas. These exhaustive information on the management of com- nurses have said that when they encounter a pa- plications, nor does it replace the need for consulta- tient who has had an ostomy, they are often un- tion with a qualified wound, ostomy, and continence sure how to care for the stoma and how to assess (WOC) nurse. -

Multiseriate Cortical Sclerenchyma Enhance Root Penetration in Compacted Soils
Multiseriate cortical sclerenchyma enhance root penetration in compacted soils Hannah M. Schneidera, Christopher F. Strocka, Meredith T. Hanlona, Dorien J. Vanheesb,c, Alden C. Perkinsa, Ishan B. Ajmeraa, Jagdeep Singh Sidhua, Sacha J. Mooneyb,d, Kathleen M. Browna, and Jonathan P. Lyncha,b,d,1 aDepartment of Plant Science, Pennsylvania State University, University Park, PA 16802; bDivision of Agricultural and Environment Sciences, School of Biosciences, University of Nottingham, Leicestershire LE12 5RD, United Kingdom; cThe James Hutton Institute, Invergowrie DD2 5DA, United Kingdom; and dCentre for Plant Integrative Biology, University of Nottingham, Leicestershire LE12 5RD, United Kingdom Edited by Philip N. Benfey, Duke University, Durham, NC, and approved January 3, 2021 (received for review June 11, 2020) Mechanical impedance limits soil exploration and resource capture Root anatomical phenes have a large effect on penetration by plant roots. We examine the role of root anatomy in regulating ability (11). Thicker roots are more resistant to buckling and plant adaptation to mechanical impedance and identify a root deflection when encountering hard soils (12, 13). However, in anatomical phene in maize (Zea mays) and wheat (Triticum aesti- maize, cortical cell wall thickness, cortical cell count, cortical cell vum ) associated with penetration of hard soil: Multiseriate cortical wall area, and stele diameter predict root penetration and bend sclerenchyma (MCS). We characterize this trait and evaluate the strength better than root diameter (14). Smaller cells in the outer utility of MCS for root penetration in compacted soils. Roots with cortical region in maize are associated with increased root pen- MCS had a greater cell wall-to-lumen ratio and a distinct UV emis- sion spectrum in outer cortical cells. -

Managing Lenticel Breakdown – Don’T Get Caught by the Snowball
Managing Lenticel Breakdown – Don’t get caught by the snowball Rob Blakey Stemilt Growers LLC, Wenatchee, WA (Formerly Washington State University, Prosser, WA) [email protected] Lenticel breakdown and other lenticel‐related disorders can be serious quality defects on apples. As with any fruit defect, it is important to correctly diagnose the defect to adapt management practices accordingly. Lenticel breakdown can be misdiagnosed as calcium burn, mild lenticel blotch pit, blister spot (Pseudomonas syringae), lenticel sunburn, bitter pit, and early stage speck rot (Phacidiopycnis washingtonensis) in Washington, etc. For diagnosis assistance, the reader is referred to the Lenticel Related Disorders Matrix from WSU (bit.ly/LenticelDisorders). Lenticel breakdown symptoms express after postharvest handling, but damage is set‐up pre‐ harvest when fruit grow rapidly and micro‐cracks develop in the fruit cuticle the epidermis and hypodermis (skin) cells are exposed to aggravation and desiccation. Symptoms are associated with lenticels because these cuticle micro‐cracks are often associated with lenticels and the lenticel provides an access point for aggravants and water egress which can eventually result in cell death and pitting around the lenticel. Aggravants may be dust, environmental protectant spray particles, salts dissolved in water, or agro‐chemicals. Lower risk fruit are generally: smaller, firmer, have less starch clearing, lower soluble solids, higher titratable acidity, lower K, Mg, and N, and higher Ca. Lenticel breakdown is most typically seen in Gala and Fuji in Washington. Lenticel breakdown is caused by a number of factors, with these factors accumulating over time (i.e. ‘snowballing’ and finally resulting in severe losses from lenticel breakdown. -

INTERXYLARY PHLOEM (Included Phloem) by Marcelo R
INTERXYLARY PHLOEM (included phloem) By Marcelo R. Pace Interxylary phloem is the presence of phloem strands embedded within the secondary xylem (wood), and produced by the activity of a single cambium (Carlquist 2013). Stems with this cambial variant are also referred to as foraminate, due to the conspicuous interxylary phloem strands in the shape of dots scattered within the wood. (Fig. 1A). However, the presence of interxylary phloem is sometimes less evident and can only be confirmed by microscopy. Fig. 1. Stem cross-section of Strychnos (Loganiaceae). A. S. guianensis, macroscopic view. B. S. millepunctata, microscopic view. Interxylary phloem can have four different ontogenetic origins. The first one is where the cambium produces phloem in both directions (inside and outside), followed by the formation of xylem only towards the inside, and as a result, enclosing the phloem in the wood. Examples of this origin are present in Thunbergia (Acanthaceae; Fig. 2A) and Dicella (Malpighiaceae; Fig 2B). However, in Thunbergia the interxylary phloem is derived from the interfascicular cambium resulting in radial patches that alternate with regions of the xylem that originate from the fascicular cambium (Fig. 2A). A second origin of the interxylary phloem is through the formation of small phloem arcs. These later become embedded in the wood through the production of xylem by the cambium on their flanks. The resulting phloem islands will contain a fragment of cambium at the bottom. This type is present in Strychnos (Loganiaceae; Fig. 1), the African species of Combretum (Combretaceae; Van Vliet, 1979), and in at least one neotropical species of Combretum (Acevedo-Rodríguez, pers. -

Tissues and Other Levels of Organization MODULE - 1 Diversity and Evolution of Life
Tissues and Other Levels of Organization MODULE - 1 Diversity and Evolution of Life 5 Notes TISSUES AND OTHER LEVELS OF ORGANIZATION You have just learnt that cell is the fundamental structural and functional unit of organisms and that bodies of organisms are made up of cells of various shapes and sizes. Groups of similar cells aggregate to collectively perform a particular function. Such groups of cells are termed “tissues”. This lesson deals with the various kinds of tissues of plants and animals. OBJECTIVES After completing this lesson, you will be able to : z define tissues; z classify plant tissues; z name the various kinds of plant tissues; z enunciate the tunica corpus theory and histogen theory; z classify animal tissues; z describe the structure and function of various kinds of epithelial tissues; z describe the structure and function of various kinds of connective tissues; z describe the structure and function of muscular tissue; z describe the structure and function of nervous tissue. 5.1 WHAT IS A TISSUE Organs such as stem, and roots in plants, and stomach, heart and lungs in animals are made up of different kinds of tissues. A tissue is a group of cells with a common origin, structure and function. Their common origin means they are derived from the same layer (details in lesson No. 20) of cells in the embryo. Being of a common origin, there are similar in structure and hence perform the same function. Several types of tissues organise to form an organ. Example : Blood, bone, and cartilage are some examples of animal tissues whereas parenchyma, collenchyma, xylem and phloem are different tissues present in the plants. -

Tree Identification: from Bark and Leaves Or Needles
Tree Identification: from bark and leaves or needles A walk in the woods can be a lot of fun, especially if you bring your kids. How do you get them to come along with you? Tell them this. “Look at the bark on the trees. Can you can find any that look like burnt potato chips, warts, cat scratches, camouflage pants or rippling muscles?” Believe it or not, these are descriptions of different kinds of tree bark. Tree ID from bark:______________________________________________________ Black cherry: The bark looks like burnt potato chips. Hackberry: The bark is bumpy and warty. Ironwood: The bark has long thin strips. With a little imagination, an ironwood can look like it is used as a scratching post by cats. They can also be easily spotted in winter because their light brown dead leaves hang on well past the first snow. Sycamore: A tree with bark that looks like camouflaged pants. The largest tree in Illinois is a sycamore, a majestic 115-foot tree near Springfield. Musclewood: a tree with smooth gray bark covering a trunk with ridges that look like they are rippling muscles. Shagbark hickory: Long strips of shaggy bark peeling at both ends. Cottonwood: Has heavy ridges that make it look like Paul Bunyan’s corduroy pants. Bur oak: Thick and gnarly bark has deep craggy furrows. The corky bark allows it to easily withstand hot forest fires. Tree ID from leaves or needles:________________________________________ Willow Oak: Has elongated leaves similar to those of a willow tree. Red pine: Red has 3 letters; a red pine has groupings of 2-3 prickly needles. -

Chapter 5: the Shoot System I: the Stem
Chapter 5 The Shoot System I: The Stem THE FUNCTIONS AND ORGANIZATION OF THE SHOOT SYSTEM PRIMARY GROWTH AND STEM ANATOMY Primary Tissues of Dicot Stems Develop from the Primary Meristems The Distribution of the Primary Vascular Bundles Depends on the Position of Leaves Primary Growth Differs in Monocot and Dicot Stems SECONDARY GROWTH AND THE ANATOMY OF WOOD Secondary Xylem and Phloem Develop from Vascular Cambium Wood Is Composed of Secondary Xylem Gymnosperm Wood Differs from Angiosperm Wood Bark Is Composed of Secondary Phloem and Periderm Buds Are Compressed Branches Waiting to Elongate Some Monocot Stems Have Secondary Growth STEM MODIFICATIONS FOR SPECIAL FUNCTIONS THE ECONOMIC VALUE OF WOODY STEMS SUMMARY ECONOMIC BOTANY: How Do You Make A Barrel? 1 KEY CONCEPTS 1. The shoot system is composed of the stem and its lateral appendages: leaves, buds, and flowers. Leaves are arranged in different patterns (phyllotaxis): alternate, opposite, whorled, and spiral. 2. Stems provide support to the leaves, buds, and flowers. They conduct water and nutrients and produce new cells in meristems (shoot apical meristem, primary and secondary meristems). 3. Dicot stems and monocot stems are usually different. Dicot stems tend to have vascular bundles distributed in a ring, whereas in monocot stems they tend to be scattered. 4. Stems are composed of the following: epidermis, cortex and pith, xylem and phloem, and periderm. 5. Secondary xylem is formed by the division of cells in the vascular cambium and is called wood. The bark is composed of all of the tissues outside the vascular cambium, including the periderm (formed from cork cambium) and the secondary phloem. -

Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris Richardii
G C A T T A C G G C A T genes Article Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii Alejandro Aragón-Raygoza 1,2 , Alejandra Vasco 3, Ikram Blilou 4, Luis Herrera-Estrella 2,5 and Alfredo Cruz-Ramírez 1,* 1 Molecular and Developmental Complexity Group at Unidad de Genómica Avanzada, Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Sede Irapuato, Km. 9.6 Libramiento Norte Carretera, Irapuato-León, Irapuato 36821, Guanajuato, Mexico; [email protected] 2 Metabolic Engineering Group, Unidad de Genómica Avanzada, Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Sede Irapuato, Km. 9.6 Libramiento Norte Carretera, Irapuato-León, Irapuato 36821, Guanajuato, Mexico; [email protected] 3 Botanical Research Institute of Texas (BRIT), Fort Worth, TX 76107-3400, USA; [email protected] 4 Laboratory of Plant Cell and Developmental Biology, Division of Biological and Environmental Sciences and Engineering (BESE), King Abdullah University of Science and Technology (KAUST), Thuwal 23955-6900, Saudi Arabia; [email protected] 5 Institute of Genomics for Crop Abiotic Stress Tolerance, Department of Plant and Soil Science, Texas Tech University, Lubbock, TX 79409, USA * Correspondence: [email protected] Received: 27 October 2020; Accepted: 26 November 2020; Published: 4 December 2020 Abstract: Ferns are a representative clade in plant evolution although underestimated in the genomic era. Ceratopteris richardii is an emergent model for developmental processes in ferns, yet a complete scheme of the different growth stages is necessary. Here, we present a developmental analysis, at the tissue and cellular levels, of the first shoot-borne root of Ceratopteris. -

Lenticels of Different Plant Species
University of Pretoria etd – Bezuidenhout, J L J (2005) CHAPTER 3 LENTICEL ONTOGENY OF ‘TOMMY ATKINS’, ‘KEITT’ AND ‘KENT’ FRUIT ABSTRACT Lenticels differentiate from existing stomata that lose their function and protrude above the fruit surface as a result of rapid anticlinal cell divisions in the epidermis of the exocarp. Based on the comparative study between different mango cultivars and mature marula fruit, it seems as if the absence of a cork cambium and cork cells in the mango lenticel could be one of the most important reasons for lenticel discolouration. An interaction between naturally occurring pigments and sap from the resin ducts in the exocarp appears to be another contributing factor for lenticel discolouration. 3.1 INTRODUCTION Lenticels can be found on the surface of stems, old roots and on several fruit types, including apples, pears, avocados and mangos (Dietz et al., 1988). In the absence of stomata will the lenticels take over the vitally important process of gaseous exchange needed for photosynthesis, respiration and transpiration (Mauseth, 1988). Postharvest discolouration of mango lenticels is a serious problem, since the resultant black markings on the fruit skin are unacceptable to consumers, consequently depreciating the economic value of the fruit (O’Hare and Prasad, 1992). The degree of lenticel discolouration may vary in different mango cultivars. In South Africa, ‘TA’ and ‘Keitt’ are two of the most important cultivars susceptible to lenticel discolouration, whereas ‘Kent’ is not known to problematic in that aspect. According to Dietz et al. (1988), mango fruit lenticels may develop from either pre-existing stomata, or from rupturing of the epidermis. -
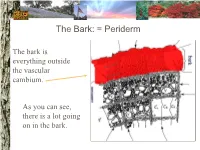
The Bark: = Periderm
The Bark: = Periderm The bark is everything outside the vascular cambium. As you can see, there is a lot going on in the bark. The Bark: periderm: phellogen (cork cambium): The phellogen is the region of cell division that forms the periderm tissues. Phellogen development influences bark appearance. The Bark: periderm: phellem (cork): Phellem replaces the epidermis as the tree increases in girth. Photosynthesis can take place in some trees both through the phellem and in fissures. The Bark: periderm: phelloderm: Phelloderm is active parenchyma tissue. Parenchyma cells can be used for storage, photosynthesis, defense, and even cell division! The Bark: phloem: Phloem tissue makes up the inner bark. However, it is vascular tissue formed from the vascular cambium. The Bark: phloem: sieve tube elements: Sieve tube elements actively transport photosynthates down the stem. Conifers have sieve cells instead. The cambium: The cambium is the primary meristem producing radial growth. It forms the phloem & xylem. The Xylem (wood): The xylem includes everything inside the vascular cambium. The Xylem: a growth increment (ring): The rings seen in many trees represent one growth increment. Growth rings provide the texture seen in wood. The Xylem: vessel elements: Hardwood species have vessel elements in addition to trachieds. Notice their location in the growth rings of this tree The Xylem: fibers: Fibers are cells with heavily lignified walls making them stiff. Many fibers in sapwood are alive at maturity and can be used for storage. The Xylem: axial parenchyma: Axial parenchyma is living tissue! Remember that parenchyma cells can be used for storage and cell division.