Chapter 5: the Shoot System I: the Stem
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Plant Terminology
PLANT TERMINOLOGY Plant terminology for the identification of plants is a necessary evil in order to be more exact, to cut down on lengthy descriptions, and of course to use the more professional texts. I have tried to keep the terminology in the database fairly simple but there is no choice in using many descriptive terms. The following slides deal with the most commonly used terms (more specialized terms are given in family descriptions where needed). Professional texts vary from fairly friendly to down-right difficult in their use of terminology. Do not be dismayed if a plant or plant part does not seem to fit any given term, or that some terms seem to be vague or have more than one definition – that’s life. In addition this subject has deep historical roots and plant terminology has evolved with the science although some authors have not. There are many texts that define and illustrate plant terminology – I use Plant Identification Terminology, An illustrated Glossary by Harris and Harris (see CREDITS) and others. Most plant books have at least some terms defined. To really begin to appreciate the diversity of plants, a good text on plant systematics or Classification is a necessity. PLANT TERMS - Typical Plant - Introduction [V. Max Brown] Plant Shoot System of Plant – stem, leaves and flowers. This is the photosynthetic part of the plant using CO2 (from the air) and light to produce food which is used by the plant and stored in the Root System. The shoot system is also the reproductive part of the plant forming flowers (highly modified leaves); however some plants also have forms of asexual reproduction The stem is composed of Nodes (points of origin for leaves and branches) and Internodes Root System of Plant – supports the plant, stores food and uptakes water and minerals used in the shoot System PLANT TERMS - Typical Perfect Flower [V. -

Transcript Profiling of a Novel Plant Meristem, the Monocot Cambium
Journal of Integrative JIPB Plant Biology Transcript profiling of a novel plant meristem, the monocot cambiumFA Matthew Zinkgraf1,2, Suzanne Gerttula1 and Andrew Groover1,3* 1. US Forest Service, Pacific Southwest Research Station, Davis, California, USA 2. Department of Computer Science, University of California, Davis, USA 3. Department of Plant Biology, University of California, Davis, USA Article *Correspondence: Andrew Groover ([email protected]) doi: 10.1111/jipb.12538 Abstract While monocots lack the ability to produce a xylem tissues of two forest tree species, Populus Research vascular cambium or woody growth, some monocot trichocarpa and Eucalyptus grandis. Monocot cambium lineages evolved a novel lateral meristem, the monocot transcript levels showed that there are extensive overlaps cambium, which supports secondary radial growth of between the regulation of monocot cambia and vascular stems. In contrast to the vascular cambium found in woody cambia. Candidate regulatory genes that vary between the angiosperm and gymnosperm species, the monocot monocot and vascular cambia were also identified, and cambium produces secondary vascular bundles, which included members of the KANADI and CLE families involved have an amphivasal organization of tracheids encircling a in polarity and cell-cell signaling, respectively. We suggest central strand of phloem. Currently there is no information that the monocot cambium may have evolved in part concerning the molecular genetic basis of the develop- through reactivation of genetic mechanisms involved in ment or evolution of the monocot cambium. Here we vascular cambium regulation. report high-quality transcriptomes for monocot cambium Edited by: Chun-Ming Liu, Institute of Crop Science, CAAS, China and early derivative tissues in two monocot genera, Yucca Received Feb. -

Tansley Review Evolution of Development of Vascular Cambia and Secondary Growth
New Phytologist Review Tansley review Evolution of development of vascular cambia and secondary growth Author for correspondence: Rachel Spicer1 and Andrew Groover2 Andrew Groover 1The Rowland Institute at Harvard, Cambridge, MA, USA; 2Institute of Forest Genetics, Pacific Tel: +1 530 759 1738 Email: [email protected] Southwest Research Station, USDA Forest Service, Davis, CA, USA Received: 29 December 2009 Accepted: 14 February 2010 Contents Summary 577 V. Evolution of development approaches for the study 587 of secondary vascular growth I. Introduction 577 VI. Conclusions 589 II. Generalized function of vascular cambia and their 578 developmental and evolutionary origins Acknowledgements 589 III. Variation in secondary vascular growth in angiosperms 581 References 589 IV. Genes and mechanisms regulating secondary vascular 584 growth and their evolutionary origins Summary New Phytologist (2010) 186: 577–592 Secondary growth from vascular cambia results in radial, woody growth of stems. doi: 10.1111/j.1469-8137.2010.03236.x The innovation of secondary vascular development during plant evolution allowed the production of novel plant forms ranging from massive forest trees to flexible, Key words: forest trees, genomics, Populus, woody lianas. We present examples of the extensive phylogenetic variation in sec- wood anatomy, wood formation. ondary vascular growth and discuss current knowledge of genes that regulate the development of vascular cambia and woody tissues. From these foundations, we propose strategies for genomics-based research in the evolution of development, which is a next logical step in the study of secondary growth. I. Introduction this pattern characterizes most extant forest trees, significant variation exists among taxa, ranging from extinct woody Secondary vascular growth provides a means of radially lycopods and horsetails with unifacial cambia (Cichan & thickening and strengthening plant axes initiated during Taylor, 1990; Willis & McElwain, 2002), to angiosperms primary, or apical growth. -

Mechanical Stress in the Inner Bark of 15 Tropical Tree Species and The
Mechanical stress in the inner bark of 15 tropical tree species and the relationship with anatomical structure Romain Lehnebach, Léopold Doumerc, Bruno Clair, Tancrède Alméras To cite this version: Romain Lehnebach, Léopold Doumerc, Bruno Clair, Tancrède Alméras. Mechanical stress in the inner bark of 15 tropical tree species and the relationship with anatomical structure. Botany / Botanique, NRC Research Press, 2019, 10.1139/cjb-2018-0224. hal-02368075 HAL Id: hal-02368075 https://hal.archives-ouvertes.fr/hal-02368075 Submitted on 18 Nov 2019 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Mechanical stress in the inner bark of 15 tropical tree species and the relationship with anatomical structure1 Romain Lehnebach, Léopold Doumerc, Bruno Clair, and Tancrède Alméras Abstract: Recent studies have shown that the inner bark is implicated in the postural control of inclined tree stems through the interaction between wood radial growth and tangential expansion of a trellis fiber network in bark. Assessing the taxonomic extent of this mechanism requires a screening of the diversity in bark anatomy and mechanical stress. The mechanical state of bark was measured in 15 tropical tree species from various botanical families on vertical mature trees, and related to the anatomical structure of the bark. -

Studies on Seed Germination, Seedling Growth, and in Vitro Shoot
HORTSCIENCE 44(3):751–756. 2009. plantlets are detached from the mother plant that are dried and planted. However, seed propagation is more feasible and recommen- Studies on Seed Germination, Seedling ded for survival of rare species (Van Wyk and Smith, 1996). If this species has to be Growth, and In Vitro Shoot Induction propagated on a large scale by means of seed or tissue culture methods, then currently there of Aloe ferox Mill., a Commercially is no basic information available on these aspects. Aloes are succulent and warm-cli- mate plants, where both temperature and Important Species water play an important role in establishing Michael W. Bairu, Manoj G. Kulkarni, Rene´e A. Street, Rofhiwa them. This study was therefore conducted to B. Mulaudzi, and Johannes Van Staden1 examine 1) the effects of different temper- atures, growth-promoting substances, and Research Centre for Plant Growth and Development, School of Biological watering frequencies on seed germination and Conservation Sciences, University of KwaZulu-Natal Pietermaritzburg, and seedling growth of A. ferox; and 2) to Private Bag X01, Scottsville 3209, South Africa assess the applicability of an in vitro propa- gation protocol developed for other Aloe spp. Additional index words. cytokinins, growth regulators, multiplication rate, smoke solutions, temperature, tissue culture Materials and Methods Abstract. A study was done to investigate the effects of some physical and chemical factors on growth and development of Aloe ferox ex vitro and in vitro. The effects of light, Seed collection. Dried seeds of A. ferox temperature, and smoke–water on seed germination, ex vitro seedling growth require- were collected between the middle to the end ments, and effect of germination medium and cytokinins on shoot induction and of August from the Botanical Garden, Uni- multiplication in vitro were investigated. -

SECONDARY GROWTH in PLANTS Compiled and Circulated by Arpita Chakraborty, Govt.Approved Part-Time Teacher, Narajole Raj College, Narajole
COMPILED AND CIRCULATED BY ARPITA CHAKRABORTY, GOVT. APPROVED PART TIME TEACHER, DEPARTMENT OF BOTANY, NARAJOLE RAJ COLLEGE. SECONDARY GROWTH IN PLANTS compiled and circulated by Arpita Chakraborty, Govt.approved Part-time teacher, Narajole Raj College, Narajole. BOTANY: SEM- IV, PAPER: GE4T:PLANT ANATOMY AND EMBRYOLOGY:UNIT-3:SECONDARY GROWTH COMPILED AND CIRCULATED BY ARPITA CHAKRABORTY, GOVT. APPROVED PART TIME TEACHER, DEPARTMENT OF BOTANY, NARAJOLE RAJ COLLEGE. •CHAPTER OUT LINE- • 1. Overview of secondary growth • 2. Growth patterns in wood and bark • 3. Commercial Uses of wood and bark BOTANY: SEM- IV, PAPER: GE4T:PLANT ANATOMY AND EMBRYOLOGY:UNIT-3:SECONDARY GROWTH COMPILED AND CIRCULATED BY ARPITA CHAKRABORTY, GOVT. APPROVED PART TIME TEACHER, DEPARTMENT OF BOTANY, NARAJOLE RAJ COLLEGE. CHAPTER OBJECTIVES- Students should have an idea of; 1. How wood and bark develop 2. How stems and roots become thicker and stronger 3. Commercial benefits of wood and bark of a plant with secondary growth BOTANY: SEM- IV, PAPER: GE4T:PLANT ANATOMY AND EMBRYOLOGY:UNIT-3:SECONDARY GROWTH COMPILED AND CIRCULATED BY ARPITA CHAKRABORTY, GOVT. APPROVED PART TIME TEACHER, DEPARTMENT OF BOTANY, NARAJOLE RAJ COLLEGE. SECONDARY GROWTH- Cambial 1.Vascular cambium a)Fusiform Initials (Vertically oriented) Secondary Xylem Secondary Phloem b)Ray Initials (Horizontally oriented) Vascular Rays Xylem rays Phloem ray 2.Cork cambium (Phellogen) Periderm Phellem (Cork cells) Phelloderm (Cork Parenchyma) BOTANY: SEM- IV, PAPER: GE4T:PLANT ANATOMY AND EMBRYOLOGY:UNIT-3:SECONDARY -
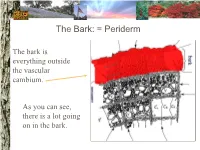
The Bark: = Periderm
The Bark: = Periderm The bark is everything outside the vascular cambium. As you can see, there is a lot going on in the bark. The Bark: periderm: phellogen (cork cambium): The phellogen is the region of cell division that forms the periderm tissues. Phellogen development influences bark appearance. The Bark: periderm: phellem (cork): Phellem replaces the epidermis as the tree increases in girth. Photosynthesis can take place in some trees both through the phellem and in fissures. The Bark: periderm: phelloderm: Phelloderm is active parenchyma tissue. Parenchyma cells can be used for storage, photosynthesis, defense, and even cell division! The Bark: phloem: Phloem tissue makes up the inner bark. However, it is vascular tissue formed from the vascular cambium. The Bark: phloem: sieve tube elements: Sieve tube elements actively transport photosynthates down the stem. Conifers have sieve cells instead. The cambium: The cambium is the primary meristem producing radial growth. It forms the phloem & xylem. The Xylem (wood): The xylem includes everything inside the vascular cambium. The Xylem: a growth increment (ring): The rings seen in many trees represent one growth increment. Growth rings provide the texture seen in wood. The Xylem: vessel elements: Hardwood species have vessel elements in addition to trachieds. Notice their location in the growth rings of this tree The Xylem: fibers: Fibers are cells with heavily lignified walls making them stiff. Many fibers in sapwood are alive at maturity and can be used for storage. The Xylem: axial parenchyma: Axial parenchyma is living tissue! Remember that parenchyma cells can be used for storage and cell division. -

Listado De Todas Las Plantas Que Tengo Fotografiadas Ordenado Por Familias Según El Sistema APG III (Última Actualización: 2 De Septiembre De 2021)
Listado de todas las plantas que tengo fotografiadas ordenado por familias según el sistema APG III (última actualización: 2 de Septiembre de 2021) GÉNERO Y ESPECIE FAMILIA SUBFAMILIA GÉNERO Y ESPECIE FAMILIA SUBFAMILIA Acanthus hungaricus Acanthaceae Acanthoideae Metarungia longistrobus Acanthaceae Acanthoideae Acanthus mollis Acanthaceae Acanthoideae Odontonema callistachyum Acanthaceae Acanthoideae Acanthus spinosus Acanthaceae Acanthoideae Odontonema cuspidatum Acanthaceae Acanthoideae Aphelandra flava Acanthaceae Acanthoideae Odontonema tubaeforme Acanthaceae Acanthoideae Aphelandra sinclairiana Acanthaceae Acanthoideae Pachystachys lutea Acanthaceae Acanthoideae Aphelandra squarrosa Acanthaceae Acanthoideae Pachystachys spicata Acanthaceae Acanthoideae Asystasia gangetica Acanthaceae Acanthoideae Peristrophe speciosa Acanthaceae Acanthoideae Barleria cristata Acanthaceae Acanthoideae Phaulopsis pulchella Acanthaceae Acanthoideae Barleria obtusa Acanthaceae Acanthoideae Pseuderanthemum carruthersii ‘Rubrum’ Acanthaceae Acanthoideae Barleria repens Acanthaceae Acanthoideae Pseuderanthemum carruthersii var. atropurpureum Acanthaceae Acanthoideae Brillantaisia lamium Acanthaceae Acanthoideae Pseuderanthemum carruthersii var. reticulatum Acanthaceae Acanthoideae Brillantaisia owariensis Acanthaceae Acanthoideae Pseuderanthemum laxiflorum Acanthaceae Acanthoideae Brillantaisia ulugurica Acanthaceae Acanthoideae Pseuderanthemum laxiflorum ‘Purple Dazzler’ Acanthaceae Acanthoideae Crossandra infundibuliformis Acanthaceae Acanthoideae Ruellia -

Secondary Tree Growth Increments 11-07
Secondary Tree Growth Increments: Ring Development & Forms by Dr. Kim D. Coder, Professor of Tree Biology & Health Care, University of Georgia Trees attempt to occupy more space and control available resources through cell divisions and cell expansion. From the end of a shoot tip to the end of a root tip, trees elongate. Along this axis of growth, trees also expand radially, which is termed “secondary growth.” Tree growth is initiated in the shoot tips (growing points / buds), root tips, vascular cambium (i.e. or simply cambium), and phellogen which generates the periderm. The first two meristems are primary generation systems and the second two are termed secondary meristems. Cambial activity and xylem formation, visible as increases in growth increment volume, is radial growth, and reviewed here. The secondary meristem phellogen, sometimes generically referred to as the cork cambium, will not be discussed here although it does increase tree diameter. Cambium The cambium is a cell generation zone which blends (within several cell thicknesses) inward into the xylem (wood) and outward into the phloem (inner bark). The cambium occupies the circumference of the wood cylinder which comprises the bulk of a tree. The cambium is responsible for modifying cell divisions, cell forms, and cell wall materials in response to mechanical, biological and defensive stresses at each point along its surface. The cambium receives signals from sensors both internally (from divid- ing and expanding cells in the cambial zone) and externally (located locally farther inside and outside the cambium, and from shoot tips and root tips.) A tree radially expands tissues as shoots and roots elongate. -

Tree Anatomy Stems and Branches
Tree Anatomy Series WSFNR14-13 Nov. 2014 COMPONENTSCOMPONENTS OFOF PERIDERMPERIDERM by Dr. Kim D. Coder, Professor of Tree Biology & Health Care Warnell School of Forestry & Natural Resources, University of Georgia Around tree roots, stems and branches is a complex tissue. This exterior tissue is the environmental face of a tree open to all sorts of site vulgarities. This most exterior of tissue provides trees with a measure of protection from a dry, oxidative, heat and cold extreme, sunlight drenched, injury ridden site. The exterior of a tree is both an ecological super highway and battle ground – comfort and terror. This exterior is unique in its attributes, development, and regeneration. Generically, this tissue surrounding a tree stem, branch and root is loosely called bark. The tissues of a tree, outside or more exterior to the xylem-containing core, are varied and complexly interwoven in a relatively small space. People tend to see and appreciate the volume and physical structure of tree wood and dismiss the remainder of stem, branch and root. In reality, tree life is focused within these more exterior thin tissue sets. Outside of the cambium are tissues which include transport cells, structural support cells, generation cells, and cells positioned to help, protect, and sustain other cells. All of this life is smeared over the circumference of a predominately dead physical structure. Outer Skin Periderm (jargon and antiquated term = bark) is the most external of tree tissues providing protection, water conservation, insulation, and environmental sensing. Periderm is a protective tissue generated over and beyond live conducting and non-conducting cells of the food transport system (phloem). -

Plant Pathology
Plant Pathology 330-1 Reading / Reference Materials CSU Extension Fact Sheets o Aspen and poplar leaf spots – #2.920 o Backyard orchard: apples and pears [pest management] – #2.800 o Backyard orchard: stone fruits [pest management] – #2.804 o Bacterial wetwood – #2.910 o Cytospora canker – #2.937 o Diseases of roses in Colorado – #2.946 o Dollar spot disease of turfgrass – #2.933 o Dutch elm disease – #5.506 o Dwarf mistletoe management – #2.925 o Fairy ring in turfgrass – #2.908 o Fire blight – #2.907 o Forest fire – Insects and diseases associated with forest fires – #6.309 o Friendly pesticides for home gardens – #2.945 o Greenhouse plant viruses (TSWV-INSV) – #2.947 o Honeylocust diseases – #2.939 o Juniper-hawthorn rust – #2.904 o Juniper-hawthorn rust – #2.904 o Leaf spot and melting out diseases – #2.909 o Necrotic ring spot in turfgrass – #2.900 o Non-chemical disease control – #2.903 o Pesticides – Friendly pesticides for home gardens – #2.945 o Pinyon pine insects and diseases – #2.948 o Powdery mildew – #2.902 o Roses – Diseases of roses in Colorado – #2.946 o Russian olive decline and gummosis – #2.942 o Strawberry diseases – #2.931 o Sycamore anthracnose – #2.930 CSU Extension Publications o Insects and diseases of woody plants of the central Rockies – 506A Curriculum developed by Mary Small, CSU Extension, Jefferson County • Colorado State University, U.S. Department of Agriculture and Colorado counties cooperating. • CSU Extension programs are available to all without discrimination. • No endorsement of products named is intended, nor is criticism implied of products not mentioned. -

Principal Types of Vegetative Shoot Apex Organization in Vascular Plants1
PRINCIPAL TYPES OF VEGETATIVE SHOOT APEX ORGANIZATION IN VASCULAR PLANTS1 RICHARD A. POPHAM Department of Botany and Plant Pathology, The Ohio State University, Columbus 10 Before progress can be made in research, a problem must be recognized. Once the problem has been perceived, a research program may be directed toward a solution. The problem of how and where a shoot grows and the organization of the shoot apex was apparently first conceived by Kaspar Friedrich Wolff (1759). Although his observations on the structure, formation, and growth of cells were fantastically inaccurate, he made a great contribution to our knowledge of the growing plant by setting forth a new and important problem. In a very real sense, Wolff is the father of developmental plant anatomy. Disagreement is the life blood of many research problems. Strenuous opposition is often engendered by a dogmatic statement or a theory which is proposed as a universal truth. Opposition to Wolff's (1759) original proposition regarding the organization and growth of shoot apices prompted plant anatomists, some 85 years later, to investigate the truth of the statement. The factual solution of the problem of shoot apex organization had its beginnings in the work of Nageli (1845). Following this work on many lower cryptogams, Nageli concluded that the cells of all tissues of the shoot of cryptogams and higher plants have their genesis in a single apical cell. The new-born apical cell theory supported by Hofmeister (1851) and others provided the impetus for a renewed, vigorous attack on the problem of shoot apex organization. A little later a new proposal, Hanstein's (1868) histogen theory was born of more careful observations and in a mind unfettered by the prevailing fanaticism of the apical cell theorists.