Internal Organisation of Plants
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Transcript Profiling of a Novel Plant Meristem, the Monocot Cambium
Journal of Integrative JIPB Plant Biology Transcript profiling of a novel plant meristem, the monocot cambiumFA Matthew Zinkgraf1,2, Suzanne Gerttula1 and Andrew Groover1,3* 1. US Forest Service, Pacific Southwest Research Station, Davis, California, USA 2. Department of Computer Science, University of California, Davis, USA 3. Department of Plant Biology, University of California, Davis, USA Article *Correspondence: Andrew Groover ([email protected]) doi: 10.1111/jipb.12538 Abstract While monocots lack the ability to produce a xylem tissues of two forest tree species, Populus Research vascular cambium or woody growth, some monocot trichocarpa and Eucalyptus grandis. Monocot cambium lineages evolved a novel lateral meristem, the monocot transcript levels showed that there are extensive overlaps cambium, which supports secondary radial growth of between the regulation of monocot cambia and vascular stems. In contrast to the vascular cambium found in woody cambia. Candidate regulatory genes that vary between the angiosperm and gymnosperm species, the monocot monocot and vascular cambia were also identified, and cambium produces secondary vascular bundles, which included members of the KANADI and CLE families involved have an amphivasal organization of tracheids encircling a in polarity and cell-cell signaling, respectively. We suggest central strand of phloem. Currently there is no information that the monocot cambium may have evolved in part concerning the molecular genetic basis of the develop- through reactivation of genetic mechanisms involved in ment or evolution of the monocot cambium. Here we vascular cambium regulation. report high-quality transcriptomes for monocot cambium Edited by: Chun-Ming Liu, Institute of Crop Science, CAAS, China and early derivative tissues in two monocot genera, Yucca Received Feb. -

PLANT ANATOMY Lecture 15 - Stele and Bundle Types
Jim Bidlack - BIO 5354/4354 PLANT ANATOMY Lecture 15 - Stele and Bundle Types I. Types of steles (stellar types); stele refers to the central vascular network if the shoot and the root (includes outermost phloem and everything to the inside of it) A. Protostele 1. No pith and usually no separate vascular bundles 2. First pattern (phylogenetically) that appeared in vascular plants 3. Predominant pattern found in roots of almost all plants and shoots of lower vascular plants 4. Types of protostele a) Haplostele - xylem is a circular mass b) Actinostele - xylem margin is not smooth; it "undulates" c) Plectostele - xylem not one mass; series of plates B. Siphonostele 1. Has a pith and can have separate vascular bundles (primary growth) 2. More advanced plants 3. Found in shoots of seed plants and in roots having a broad stele (monocots) 4. Types of siphonostele a) Amphiphloic - phloem found on both sides of the xylem 1) Solenostele - very few or little leaf gaps widely separated (continuous cylinder) 2) Dictyostele - leaf gaps are abundant b) Ectophloic - phloem found only to outside of the xylem 1) Eustele (dicots) - one ring of vascular bundles surround a pith 2) Atactostele (monocots) - scattered bundle arrangement II. Stellar patterns at the node (Nodal pattern) A. Different because this is where leaves and buds are attached B. Area above & behind the leaf or bud is the leaf gap C. Area showing where the leaf (bundle(s)) attaches to stem is called leaf trace D. Examples of patterns: 1. Unilacunar, 1 trace; unilacunar, 3 trace; trilacunar, 3 trace III. -

Tansley Review Evolution of Development of Vascular Cambia and Secondary Growth
New Phytologist Review Tansley review Evolution of development of vascular cambia and secondary growth Author for correspondence: Rachel Spicer1 and Andrew Groover2 Andrew Groover 1The Rowland Institute at Harvard, Cambridge, MA, USA; 2Institute of Forest Genetics, Pacific Tel: +1 530 759 1738 Email: [email protected] Southwest Research Station, USDA Forest Service, Davis, CA, USA Received: 29 December 2009 Accepted: 14 February 2010 Contents Summary 577 V. Evolution of development approaches for the study 587 of secondary vascular growth I. Introduction 577 VI. Conclusions 589 II. Generalized function of vascular cambia and their 578 developmental and evolutionary origins Acknowledgements 589 III. Variation in secondary vascular growth in angiosperms 581 References 589 IV. Genes and mechanisms regulating secondary vascular 584 growth and their evolutionary origins Summary New Phytologist (2010) 186: 577–592 Secondary growth from vascular cambia results in radial, woody growth of stems. doi: 10.1111/j.1469-8137.2010.03236.x The innovation of secondary vascular development during plant evolution allowed the production of novel plant forms ranging from massive forest trees to flexible, Key words: forest trees, genomics, Populus, woody lianas. We present examples of the extensive phylogenetic variation in sec- wood anatomy, wood formation. ondary vascular growth and discuss current knowledge of genes that regulate the development of vascular cambia and woody tissues. From these foundations, we propose strategies for genomics-based research in the evolution of development, which is a next logical step in the study of secondary growth. I. Introduction this pattern characterizes most extant forest trees, significant variation exists among taxa, ranging from extinct woody Secondary vascular growth provides a means of radially lycopods and horsetails with unifacial cambia (Cichan & thickening and strengthening plant axes initiated during Taylor, 1990; Willis & McElwain, 2002), to angiosperms primary, or apical growth. -

Plant Water Relations: Absorption, Transport and Control Mechanisms
5 Plant Water Relations: Absorption, Transport and Control Mechanisms Geraldo Chavarria1 and Henrique Pessoa dos Santos2 1The University of Passo Fundo 2Embrapa Grape & Wine Brazil 1. Introduction Although water is abundant on Earth - covering 71% of the total surface - its distribution is not uniform and can easily cause restrictions in availability to vegetal production. At global scale, these restrictions are easily observed in dry climates and can appear in other regions which do not currently experience drought, as provided by the future backdrop of climate change (IPCC, 2007). The influences of water restriction on losses in the production and distribution of vegetation on the terrestrial surface are significantly larger than all other losses combined which are caused by biotic and abiotic factors (Boyer, 1985). This striking effect of water on plants emerges from its physiological importance, being an essential factor for successful plant growth, involving photosynthesis and several other biochemical processes such as the synthesis of energetic composites and new tissue. Therefore, in order to characterise the growth and productive behaviour of plant species it is essential to have an understanding of plant water relations, as well as the consequences of an inadequate water supply. Broadly, the water state of a plant is controlled by relative rates of loss and absorption, moreover it depends on the ability to adjust and keep an adequate water status. This will be considered throughout this chapter. 2. Absorption and water flow through plants Independent of the species, plants require from the soil a water volume that overcomes its metabolic necessities. Through the transpiration process plants transmit to the atmosphere the majority of the water absorbed from soil (generally around 90%). -

Anatomy of Leaf Apical Hydathodes in Four Monocotyledon Plants of Economic and Academic Relevance Alain Jauneau, Aude Cerutti, Marie-Christine Auriac, Laurent D
Anatomy of leaf apical hydathodes in four monocotyledon plants of economic and academic relevance Alain Jauneau, Aude Cerutti, Marie-Christine Auriac, Laurent D. Noël To cite this version: Alain Jauneau, Aude Cerutti, Marie-Christine Auriac, Laurent D. Noël. Anatomy of leaf apical hydathodes in four monocotyledon plants of economic and academic relevance. PLoS ONE, Public Library of Science, 2020, 15 (9), pp.e0232566. 10.1371/journal.pone.0232566. hal-02972304 HAL Id: hal-02972304 https://hal.inrae.fr/hal-02972304 Submitted on 20 Oct 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License PLOS ONE RESEARCH ARTICLE Anatomy of leaf apical hydathodes in four monocotyledon plants of economic and academic relevance 1☯ 2☯ 1,2 2 Alain Jauneau *, Aude Cerutti , Marie-Christine Auriac , Laurent D. NoeÈlID * 1 FeÂdeÂration de Recherche 3450, Universite de Toulouse, CNRS, Universite Paul Sabatier, Castanet- Tolosan, France, 2 LIPM, Universite de Toulouse, INRAE, CNRS, Universite Paul Sabatier, Castanet- Tolosan, France ☯ These authors contributed equally to this work. a1111111111 * [email protected] (AJ); [email protected] (LN) a1111111111 a1111111111 a1111111111 a1111111111 Abstract Hydathode is a plant organ responsible for guttation in vascular plants, i.e. -

Chapter 5: the Shoot System I: the Stem
Chapter 5 The Shoot System I: The Stem THE FUNCTIONS AND ORGANIZATION OF THE SHOOT SYSTEM PRIMARY GROWTH AND STEM ANATOMY Primary Tissues of Dicot Stems Develop from the Primary Meristems The Distribution of the Primary Vascular Bundles Depends on the Position of Leaves Primary Growth Differs in Monocot and Dicot Stems SECONDARY GROWTH AND THE ANATOMY OF WOOD Secondary Xylem and Phloem Develop from Vascular Cambium Wood Is Composed of Secondary Xylem Gymnosperm Wood Differs from Angiosperm Wood Bark Is Composed of Secondary Phloem and Periderm Buds Are Compressed Branches Waiting to Elongate Some Monocot Stems Have Secondary Growth STEM MODIFICATIONS FOR SPECIAL FUNCTIONS THE ECONOMIC VALUE OF WOODY STEMS SUMMARY ECONOMIC BOTANY: How Do You Make A Barrel? 1 KEY CONCEPTS 1. The shoot system is composed of the stem and its lateral appendages: leaves, buds, and flowers. Leaves are arranged in different patterns (phyllotaxis): alternate, opposite, whorled, and spiral. 2. Stems provide support to the leaves, buds, and flowers. They conduct water and nutrients and produce new cells in meristems (shoot apical meristem, primary and secondary meristems). 3. Dicot stems and monocot stems are usually different. Dicot stems tend to have vascular bundles distributed in a ring, whereas in monocot stems they tend to be scattered. 4. Stems are composed of the following: epidermis, cortex and pith, xylem and phloem, and periderm. 5. Secondary xylem is formed by the division of cells in the vascular cambium and is called wood. The bark is composed of all of the tissues outside the vascular cambium, including the periderm (formed from cork cambium) and the secondary phloem. -

3115 SAPS Operating Instructions
OPERATING INSTRUCTIONS FIELD PLANT WATER STATUS CONSOLE March 2017 Fig. 1 - Model 3115 Portable Plant Water Console shown with 22 c/f Compressed Gas Cylinder, 3072V22 SOILMOISTURE EQUIPMENT CORP. P.O. Box 30025, Santa Barbara, CA 93105 U.S.A. Telephone 805-964-3525 - Fax No. 805-683-2189 Email: [email protected] - Website: http://www.soilmoisture.com Table of Contents Chapter - Page 1 Description ............................................................................................................................... 1 - 4 2 Technical Specification ............................................................................................................ 2 - 5 2.1 Weight .......................................................................................................................... 2 - 5 2.2 Dimensions ................................................................................................................... 2 - 5 2.3 Pressure Vessel ............................................................................................................. 2 - 5 2.4 Gauges .......................................................................................................................... 2 - 5 2.5 Valves ........................................................................................................................... 2 - 5 2.6 Connecting Hose .......................................................................................................... 2 - 5 2.7 Pressure Tank .............................................................................................................. -
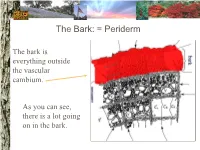
The Bark: = Periderm
The Bark: = Periderm The bark is everything outside the vascular cambium. As you can see, there is a lot going on in the bark. The Bark: periderm: phellogen (cork cambium): The phellogen is the region of cell division that forms the periderm tissues. Phellogen development influences bark appearance. The Bark: periderm: phellem (cork): Phellem replaces the epidermis as the tree increases in girth. Photosynthesis can take place in some trees both through the phellem and in fissures. The Bark: periderm: phelloderm: Phelloderm is active parenchyma tissue. Parenchyma cells can be used for storage, photosynthesis, defense, and even cell division! The Bark: phloem: Phloem tissue makes up the inner bark. However, it is vascular tissue formed from the vascular cambium. The Bark: phloem: sieve tube elements: Sieve tube elements actively transport photosynthates down the stem. Conifers have sieve cells instead. The cambium: The cambium is the primary meristem producing radial growth. It forms the phloem & xylem. The Xylem (wood): The xylem includes everything inside the vascular cambium. The Xylem: a growth increment (ring): The rings seen in many trees represent one growth increment. Growth rings provide the texture seen in wood. The Xylem: vessel elements: Hardwood species have vessel elements in addition to trachieds. Notice their location in the growth rings of this tree The Xylem: fibers: Fibers are cells with heavily lignified walls making them stiff. Many fibers in sapwood are alive at maturity and can be used for storage. The Xylem: axial parenchyma: Axial parenchyma is living tissue! Remember that parenchyma cells can be used for storage and cell division. -

Effect of Stele Type in the Growth Rotation of Dalbergia Melanoxylon
International Journal of Plant and Forestry Sciences Vol. 2, No. 3, May 2015, pp. 1 -10 Available online at http://ijpfs.com/ Research article Effect of Stele type in the growth rotation of Dalbergia melanoxylon. Dr. Washa B. Washa Mkwawa University College of Education Private Bag, MUCE Iringa Tanzania. E-mail: [email protected], +255 752 356 709 This work is licensed under a Creative Commons Attribution 4.0 International License. _____________________________________________________________________________________________ Abstract Examination of the effect of stele type in Dalbergia melanoxylon growth was conducted. Examination of stems to observe stele type and tissue water potential of Barreveld Faux, Cupressus sempervirens and Dalbergia melanoxylon was carried between 2nd – 23rd April 2015 at Mkwawa University Laboratories. About 120 stained stem sections were examined for stele type while other 120 stem pieces were incubated in Nacl solution for water potential. Forty (40) stained sections indicated ectophloic siphonostele in Barreveld Faux and C. sempervirens while 40 sections of D. melanoxylon indicated atactostele. Calculated water potential of Barreveld Faux was -0.01158597 bars and that of C. sempervirens was -0.01257201 bars while that of D. melanoxylon was -0.00320463 bars. Results found that, low water potential in D. melanoxylon is a factor for its slow growth rotation and this is due to the atactostele type existing in D. melanoxylon. A hard Blackwood and valued wood in D. melanoxylon is brought by this stele type. In order to initiate rapid growth rotation in D. melanoxylon is recommended to conduct genetic recombination of the D. melanoxylon with other species which have an easily growing wood although this can lower the hardwood quality and value of the D. -

Tree Anatomy Stems and Branches
Tree Anatomy Series WSFNR14-13 Nov. 2014 COMPONENTSCOMPONENTS OFOF PERIDERMPERIDERM by Dr. Kim D. Coder, Professor of Tree Biology & Health Care Warnell School of Forestry & Natural Resources, University of Georgia Around tree roots, stems and branches is a complex tissue. This exterior tissue is the environmental face of a tree open to all sorts of site vulgarities. This most exterior of tissue provides trees with a measure of protection from a dry, oxidative, heat and cold extreme, sunlight drenched, injury ridden site. The exterior of a tree is both an ecological super highway and battle ground – comfort and terror. This exterior is unique in its attributes, development, and regeneration. Generically, this tissue surrounding a tree stem, branch and root is loosely called bark. The tissues of a tree, outside or more exterior to the xylem-containing core, are varied and complexly interwoven in a relatively small space. People tend to see and appreciate the volume and physical structure of tree wood and dismiss the remainder of stem, branch and root. In reality, tree life is focused within these more exterior thin tissue sets. Outside of the cambium are tissues which include transport cells, structural support cells, generation cells, and cells positioned to help, protect, and sustain other cells. All of this life is smeared over the circumference of a predominately dead physical structure. Outer Skin Periderm (jargon and antiquated term = bark) is the most external of tree tissues providing protection, water conservation, insulation, and environmental sensing. Periderm is a protective tissue generated over and beyond live conducting and non-conducting cells of the food transport system (phloem). -

Stelar Evolution
Stelar System of Plant: Definition and Types Definition of Stelar System: According to the older botanists, the vascular bundle is the fundamental unit in the vascular system of pteridophytes and higher plants. Van Tieghem and Douliot (1886) interpreted the plant body of vascular plant in the different way. According to them, the fundamental parts of a shoot are the cortex and a central cylinder, is known as stele. Thus the stele is defined as a central vascular cylinder, with or without pith and delimited the cortex by endodermis. The term stele has been derived from a Greek word meaning pillar. Van Tieghem and Douliot (1886) recognized only three types of steles. They also thought that the monostelic shoot were rare in comparison of polystelic shoots. It is an established fact that all shoots are monostelic and polystelic condition rarely occurs. The stele of the stem remains connected with that of leaf by a vascular connection known as the leaf supply. Types of Steles: 1. Protostele: Jeffrey (1898), for the first time pointed out the stelar theory from the point of view of the phylogeny. According to him, the primitive type of stele is protostele. In protostele, the vascular tissue is a solid mass and the central core of the xylem is completely surrounded by the strand of phloem. This is the most primitive and simplest type of stele. There are several forms of protostele: (a) Haplostele: This is the most primitive type of protostele. Here the central solid smooth core of xylem remains surrounded by phloem (e.g., in Selaginella spp.). -

Differences Between Transpiration and Guttation
BIOLOGY TRANSPIRATION Significance and Factors affecting Transpiration Factors Affecting the Rate of Transpiration •On a bright sunny day, stomata open fully, so transpiration is increased. •On a cloudy day, stomata open partially, so Intensity of sunlight transpiration is reduced. •At night, stomata close; hence, transpiration is greatly reduced or negligible. •Increase in temperature of the air increases the rate Temperature of transpiration. •Transpiration increases with rapid or active air Velocity of wind movement. Humidity •If the air is humid, the rate of transpiration is reduced. •Increase in the CO2 level in the atmosphere over Carbon dioxide normal 0.03% causes stomatal closure. Hence, it decreases the rate of transpiration. •With decrease in atmospheric pressure, the rate of Atmospheric pressure transpiration increases. www.topperlearning.com 2 BIOLOGY TRANSPIRATION Adaptation in Plants to Control Excessive Transpiration Plants which grow in dry climate have evolved a variety of adaptations to curtail transpiration. Morphological Leaves may be modified into spines as in cactus or into needles as in pines. Adaptations Spines Needles Leaves may be folded or rolled up. Rolled up Leaves Leaves may be shed. Example: deciduous trees. Deciduous Trees Anatomical The number of stomata is reduced, and they may be sunken in pits. Adaptations Sunken Stoma www.topperlearning.com 3 BIOLOGY TRANSPIRATION Structure of Sunken Stomata A thick waxy cuticle develops on the leaves. Example: Banyan tree, evergreen trees. Banyan Tree Shrubs and grass develop a waterproof covering of cork or bark. A multiple epidermis may develop in some leaves. www.topperlearning.com 4 BIOLOGY TRANSPIRATION Significance of Transpiration Cooling Effect Suction Force Distribution of Water and Mineral Salts Evaporation reduces the As water evaporates from leaves, Higher the rate of transpiration, temperature of leaf a suction force is created.