Almost Reason Enough for Having Eyes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
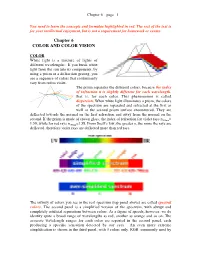
Chapter 6 COLOR and COLOR VISION
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment. -

Color Vision Mechanisms
11 COLOR VISION MECHANISMS Andrew Stockman Department of Visual Neuroscience UCL Institute of Opthalmology London, United KIngdom David H. Brainard Department of Psychology University of Pennsylvania Philadelphia, Pennsylvania 11.1 GLOSSARY Achromatic mechanism. Hypothetical psychophysical mechanisms, sometimes equated with the luminance mechanism, which respond primarily to changes in intensity. Note that achromatic mech- anisms may have spectrally opponent inputs, in addition to their primary nonopponent inputs. Bezold-Brücke hue shift. The shift in the hue of a stimulus toward either the yellow or blue invariant hues with increasing intensity. Bipolar mechanism. A mechanism, the response of which has two mutually exclusive types of out- put that depend on the balance between its two opposing inputs. Its response is nulled when its two inputs are balanced. Brightness. A perceptual measure of the apparent intensity of lights. Distinct from luminance in the sense that lights that appear equally bright are not necessarily of equal luminance. Cardinal directions. Stimulus directions in a three-dimensional color space that silence two of the three “cardinal mechanisms.” These are the isolating directions for the L+M, L–M, and S–(L+M) mech- anisms. Note that the isolating directions do not necessarily correspond to mechanism directions. Cardinal mechanisms. The second-site bipolar L–M and S–(L+M) chromatic mechanisms and the L+M luminance mechanism. Chromatic discrimination. Discrimination of a chromatic target from another target or back- ground, typically measured at equiluminance. Chromatic mechanism. Hypothetical psychophysical mechanisms that respond to chromatic stimuli, that is, to stimuli modulated at equiluminance. Color appearance. Subjective appearance of the hue, brightness, and saturation of objects or lights. -

Color Vision
Color Vision Lecture 10 Chapter 5, Part A Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Princeton University, Fall 2017 1 Exam #1: Thursday 10/19 Format: multiple-choice, fill-in-the-blank, & short answer What to study: - all material from lectures & slides - precept readings (basic gist & findings of each article) (If something appeared only in the book, and not at all in class or precept or slides, you can probably safely ignore it) Review session: tonight @ 7:00 in PNI A30 2 2015 called…. Grey-and-green, or pink-and-white? 3 4 5 • color vision has evolutionary value • lack of color vision ≠ black & white 6 Basic Principles of Color Perception The book says: “Color is not a physical property but a psychophysical property” 7 Basic Principles of Color Perception • Most of the light we see is reflected • Typical light sources: Sun, light bulb, LED screen • We see only part of the electromagnetic spectrum(between 400 and 700 nm). Why?? 8 Basic Principles of Color Perception • Why only 400-700 nm? (Pomerantz, Rice U.) Suggestion: unique ability to penetrate sea water 9 Basic Principles of Color Perception Q: How many numbers would you need to write down to specify the spectral properties of a light source? A: It depends on how you “bin” up the spectrum • One number for each spectral “bin”: 20 17 16 15 13 12 10 example: 13 5 bins energy 0 0 0 0 0 10 Basic Principles of Color Perception Device: hyper-spectral camera - measures the amount of energy (or number of photons) in each small range of wavelengths - can use thousands of bins (or “frequency bands”) instead of just the 13 shown here 20 17 16 15 13 12 10 5 energy 0 0 0 0 0 11 Basic Principles of Color Perception Some terminology for colored light: spectral - referring to the wavelength of light the illuminant - light source power spectrum - this curve. -

Color Vision and Night Vision Chapter Dingcai Cao 10
Retinal Diagnostics Section 2 For additional online content visit http://www.expertconsult.com Color Vision and Night Vision Chapter Dingcai Cao 10 OVERVIEW ROD AND CONE FUNCTIONS Day vision and night vision are two separate modes of visual Differences in the anatomy and physiology (see Chapters 4, perception and the visual system shifts from one mode to the Autofluorescence imaging, and 9, Diagnostic ophthalmic ultra- other based on ambient light levels. Each mode is primarily sound) of the rod and cone systems underlie different visual mediated by one of two photoreceptor classes in the retina, i.e., functions and modes of visual perception. The rod photorecep- cones and rods. In day vision, visual perception is primarily tors are responsible for our exquisite sensitivity to light, operat- cone-mediated and perceptions are chromatic. In other words, ing over a 108 (100 millionfold) range of illumination from near color vision is present in the light levels of daytime. In night total darkness to daylight. Cones operate over a 1011 range of vision, visual perception is rod-mediated and perceptions are illumination, from moonlit night light levels to light levels that principally achromatic. Under dim illuminations, there is no are so high they bleach virtually all photopigments in the cones. obvious color vision and visual perceptions are graded varia- Together the rods and cones function over a 1014 range of illu- tions of light and dark. Historically, color vision has been studied mination. Depending on the relative activity of rods and cones, as the salient feature of day vision and there has been emphasis a light level can be characterized as photopic (cones alone on analysis of cone activities in color vision. -

Color Appearance Models Second Edition
Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Color Appearance Models Wiley–IS&T Series in Imaging Science and Technology Series Editor: Michael A. Kriss Formerly of the Eastman Kodak Research Laboratories and the University of Rochester The Reproduction of Colour (6th Edition) R. W. G. Hunt Color Appearance Models (2nd Edition) Mark D. Fairchild Published in Association with the Society for Imaging Science and Technology Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratory Rochester Institute of Technology, USA Copyright © 2005 John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England Telephone (+44) 1243 779777 This book was previously publisher by Pearson Education, Inc Email (for orders and customer service enquiries): [email protected] Visit our Home Page on www.wileyeurope.com or www.wiley.com All Rights Reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, scanning or otherwise, except under the terms of the Copyright, Designs and Patents Act 1988 or under the terms of a licence issued by the Copyright Licensing Agency Ltd, 90 Tottenham Court Road, London W1T 4LP, UK, without the permission in writing of the Publisher. Requests to the Publisher should be addressed to the Permissions Department, John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Sussex PO19 8SQ, England, or emailed to [email protected], or faxed to (+44) 1243 770571. This publication is designed to offer Authors the opportunity to publish accurate and authoritative information in regard to the subject matter covered. -

Human Color Vision and the Unsaturated Blue Color of the Daytime Sky Glenn S
Human color vision and the unsaturated blue color of the daytime sky Glenn S. Smitha) School of Electrical and Computer Engineering, Georgia Institute of Technology, Atlanta, Georgia 30332-0250 ͑Received 21 July 2004; accepted 10 December 2004͒ The usual answer to the question ‘‘Why is the sky blue?’’ is based only on Rayleigh scattering from the molecules in the atmosphere and makes little mention of the contribution of color vision. We supplement this answer with a quantitative discussion of the role color vision plays in determining the appearance of the daytime sky. The anatomy of the human eye is reviewed, and its response as a function of wavelength is described via the spectral sensitivities of the cones. Color matching is examined for a mixture of monochromatic lights and for the spectrum of the daytime sky. The spectral irradiance of skylight is shown to be a metameric match to unsaturated blue light. A simple experiment is described suitable for classroom use or a student project. © 2005 American Association of Physics Teachers. ͓DOI: 10.1119/1.1858479͔ I. INTRODUCTION lengths, the measurements fall below the theory. The conse- quences of this difference will be discussed in more detail in The question ‘‘Why is the sky blue?’’ is posed and an- Sec. III. swered in physics textbooks at all levels, from the introduc- As a guide, we have indicated at the bottom of Fig. 1͑b͒ tory, such as Serway and Jewett,1 to the more advanced texts 2 the color that the average observer associates with mono- on electromagnetism, such as Jackson. The popularity of chromatic light at a given wavelength, for example, blue for this topic rests on the fact that students appreciate a physical wavelengths around 460 nm.8 Clearly, the scattered radiation explanation for such an easily observed and beautiful natural is greatest at the shorter wavelengths, the blue end of the phenomenon. -

Color Vision Testing
Color Vision Testing Lea Color Vision Test The stimulus size can be reduced by using a dark gray restriction ring with an opening of 1.2 cm (.47 in) in diameter. To The Tester The large stimulus area corresponds to the visual angle of 3.8° when testing at 50 cm (20 in) and to 6.3° when testing at 30 cm (12 in). 1155 Jansen Farm Drive Quantitative measurement of color vision is an important diagnostic The small stimulus is seen as the recommended 1.5° stimulus at a Elgin, IL 60123 test used to define the degree of hereditary color vision defects found distance of 46 cm (18 in). When testing young children or persons Phone: 800-362-3860 Fax: 888-362-2576 in screening with pseudoisochromatic tests and in evaluating defi- with low vision, the distance is often much shorter than 30 cm, thus Phone: 847-841-1145 Fax: 847-841-1149 cient color vision from acquired disorders. the size of the large stimulus becomes 9.5° at 20 cm (8 in) and 19° www.good-lite.com The Panel 16 Quantitative Color Vision Test is unique from other at 10 cm (4 in). quantitative color vision tests because it uses large cap sizes, which The color surface has a protective coating, which decreases the risk gives more information about color vision function both in normally of the stimulus area getting smudged1. sighted and low vision individuals. The Panel 16 Color Vision Test consists of a set of a “pilot” of “pilots” Color Vision and 15 test caps of the same hues as in the Farnsworth Panel D-15 Neural Functions in Color Vision Test. -

Color Perception and Environmentally Based Impairments
Encyclopedia of Color Science and Technology DOI 10.1007/978-3-642-27851-8_71-5 # Springer Science+Business Media New York 2013 Color Perception and Environmentally Based Impairments Galina V. Paramei* Department of Psychology, Liverpool Hope University, Liverpool, UK Synonyms Acquired color vision impairment; Acquired color vision loss; Dyschromatopsia Definition Decreased discrimination of colors caused by adverse environment, such as long-term occupational exposure to or consumption of drugs, substances, and food containing neurotoxic chemicals. Color vision early manifests adverse effects of exposure to an environment that contains neuro- toxic substances [1, 2]. The acquired color vision impairments, or dyschromatopsias, can be very subtle (subclinical) but also may vary considerably in severity, increasing or decreasing as long as the responsible agent persists, and can become irreversible under long-term exposure and/or agent dose. There are several scenarios of exposure to hazardous chemical agents in the environment: i. Long-term occupational exposure to certain substances (e.g., neurotoxic metals, organic solvents, carbon disulfide, etc.) ii. Self-administered chronic consumption of substances containing neurotoxic chemicals (e.g., alcohol, tobacco) iii. Side effects from pharmacological treatment of medical conditions (e.g., cardiovascular, antiepileptic, or antituberculosis drugs) iv. Consumption of food contaminated by neurotoxic elements through the food chain (e.g., mercury) General Characteristics of Neurotoxin-Induced Color Vision Impairments Acquired color vision defects, unlike congenital ones, are noticeable to the observer: recently affected subjects name the stimuli as they see them – in contrast to subjects with congenital color vision defects where there is compensatory adaption of their color naming to that of normal trichromats. -

Color Appearance Models Second Edition
Color Appearance Models Second Edition Mark D. Fairchild Munsell Color Science Laboratoiy Rochester Institute of Technology, USA John Wiley & Sons, Ltd Contents Series Preface xiii Preface XV Introduction xix 1 Human Color Vision 1 1.1 Optics of the Eye 1 1.2 The Retina 6 1.3 Visual Signal Processing 12 1.4 Mechanisms of Color Vision 17 1.5 Spatial and Temporal Properties of Color Vision 26 1.6 Color Vision Deficiencies 30 1.7 Key Features for Color Appearance Modeling 34 2 Psychophysics 35 2.1 Psychophysics Defined 36 2.2 Historical Context 37 2.3 Hierarchy of Scales 40 2.4 Threshold Techniques 42 2.5 Matching Techniques 45 2.6 One-Dimensional Scaling 46 2.7 Multidimensional Scaling 49 2.8 Design of Psychophysical Experiments 50 2.9 Importance in Color Appearance Modeling 52 3 Colorimetry 53 3.1 Basic and Advanced Colorimetry 53 3.2 Whyis Color? 54 3.3 Light Sources and Illuminants 55 3.4 Colored Materials 59 3.5 The Human Visual Response 66 3.6 Tristimulus Values and Color Matching Functions 70 3.7 Chromaticity Diagrams 77 3.8 CIE Color Spaces 78 3.9 Color Difference Specification 80 3.10 The Next Step , 82 viii CONTENTS 4 Color Appearance Terminology 83 4.1 Importance of Definitions 83 4.2 Color 84 4.3 Hue 85 4.4 Brightness and Lightness 86 4.5 Colorfulness and Chroma 87 4.6 Saturation 88 4.7 Unrelated and Related Colors 88 4.8 Definitions in Equations 90 4.9 Brightness-Colorfulness vs Lightness-Chroma 91 5 Color Order Systems 94 5.1 Overvlew and Requirements 94 5.2 The Munsell Book of Color 96 5.3 The Swedish Natural Color System (NCS)