Color Vision and Night Vision Chapter Dingcai Cao 10
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

How Can Retroreflective Clothing Provide More Safety Through Visibility in a Semi-Dark Urban Environment? a Study Taking Plac
MASTER’S THESIS How can retroreflective clothing BY VIOLA SCHMITZ provide more safety through visibility in a semi-dark urban Royal Institute of Technology environment? KTH School of Architecture Master’s Program in A study taking place in Scandinavia. Architectural Lighting Design 2018-2019 24.05.2019 AF270X VT19-1 Tutor: Foteini Kyriakidou 0 Index Abstract P. 2 1. Introduction P. 2 2. Background P. 3 2.1. Urban Background P. 4 2.2. Biological background P. 4 2.2.1. Reflexes and reactions P. 4 2.2.2. Types of vision P. 4 2.2.3. Effect of pattern P. 5 recognition 2.2.4. Human field of vision P. 5 3. Analysis P. 6 3.1. Analysis: Retroreflectors P. 6 3.2. Analysis: Existing products P. 7 4. Methodology P. 9 5. Methods P. 10 5.1. Survey: P. 10 Lines defining the human body 5.2. Video Experiment: P. 10 Designs in motion 5.2.1. Analysis: Location P. 10 5.2.2. Video Experiment P. 11 5.2.3. Procedure P. 12 5.3. Experimental survey: P. 12 Size of a human 5.4. Visualization: P. 13 Pattern recognition in surroundings 6. Results P. 14 6.1. Survey: P. 14 Lines defining the human body 6.2. Video Experiment: P. 15 Designs in motion 6.2.1. Analysis: Location P. 15 6.2.2. Video Experiment P. 16 6.2.3. Observation P. 17 6.3. Experimental survey: P. 17 Size of a human 6.4. Visualization: Pattern P. 17 recognition in surroundings 7. Discussion P. -

Visual Purple and the Photopic Luminosity Curve
Br J Ophthalmol: first published as 10.1136/bjo.32.11.793 on 1 November 1948. Downloaded from THE BRITISH JOURNAL OF OPHTHALMOLOGY NOVEMBER, 1948 by copyright. COMMUNICATIONS VISUAL PURPLE AND THE PHOTOPIC LUMINOSITY- CURVE* BY H. J. A. DARTNALL http://bjo.bmj.com/ FROM THE VISION RESEARCH UNIT, 'MEDICAL RESEARCH COUNCIL, INSTITUTE OF OPHTHALMOLOGY, LONDON Introduction THE precise relationship which has been established between the scotopic luminosity curve a'nd visual purple (Dartnall & Goodeve, 1937; Wald, 19138) leads one to expect that the photopic luminositv on September 26, 2021 by guest. Protected curve is'similarly related to some " photopic " pigment or pig- ments. This latter hypothesis has given rise to considerable speculation, particularly since the evidence for the existence of retinal pigments, other than the various forms of visual purple, is not unequivocal. In any case there is no evidence for such additional pigments in the human retina. Visual purple mediates scotopic- vision by virtue of the photo- chemical changes it undergoes on exposure to light. The rate of * Received for Publication, July 12, 1948. Br J Ophthalmol: first published as 10.1136/bjo.32.11.793 on 1 November 1948. Downloaded from 794 H. J. A. DARTNALL any photochemical change is gQverned by the value of the product y Ia where y is the quantqm efficiency of the process and Ia the intensity of the absorbed light expressed in quanta per second. Since the eye in the photopic condition is much less sensitive than in the scotopic state it follows that the value of y la must be correspondingly smaller for the photopic process. -

The Purkinje Rod-Cone Shift As a Function of Luminance and Retinal Eccentricity
Vision Research 42 (2002) 2485–2491 www.elsevier.com/locate/visres Rapid communication The Purkinje rod-cone shift as a function of luminance and retinal eccentricity Stuart Anstis * Department of Psychology, University of California, San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0109, USA Received 14 September 2000; received in revised form 25 June 2002 Abstract In the Purkinje shift, the dark adapted eye becomes more sensitive to blue than to red as the retinal rods take over from the cones. A striking demonstration of the Purkinje shift, suitable for classroom use, is described in which a small change in viewing distance can reverse the perceived direction of a rotating annulus. We measured this shift with a minimum-motion stimulus (Anstis & Cavanagh, Color Vision: Physiology & Psychophysics, Academic Press, London, 1983) that converts apparent lightness of blue versus red into apparent motion. We filled an iso-eccentric annulus with radial red/blue sectors, and arranged that if the blue sectors looked darker (lighter) than the red sectors, the annulus would appear to rotate to the left (right). At equiluminance the motion appeared to vanish. Our observers established these motion null points while viewing the pattern at various retinal eccentricities through various neutral density filters. Results: The luminous efficiency of blue (relative to red) increased linearly with eccentricity at all adaptation levels, and the more the dark-adaptation, the steeper the slope of the eccentricity function. Thus blue sensitivity was a linear function of eccentricity and an exponential function of filter factor. Blue sensitivity increased linearly with eccentricity, and each additional log10 unit of dark adaptation changed the slope threefold. -
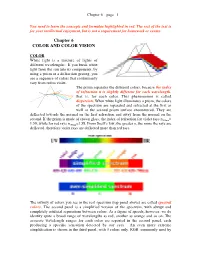
Chapter 6 COLOR and COLOR VISION
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs
sustainability Article Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs Enrique Navarrete-de Galvez 1 , Alfonso Gago-Calderon 1,* , Luz Garcia-Ceballos 2, Miguel Angel Contreras-Lopez 2 and Jose Ramon Andres-Diaz 1 1 Proyectos de Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (E.N.-d.G.); [email protected] (J.R.A.-D.) 2 Expresión Gráfica en la Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (L.G.-C.); [email protected] (M.A.C.-L.) * Correspondence: [email protected]; Tel.: +34-951-952-268 Abstract: The sensitivity of the human eye varies with the different lighting conditions to which it is exposed. The cone photoreceptors perceive the color and work for illuminance conditions greater than 3.00 cd/m2 (photopic vision). Below 0.01 cd/m2, the rods are the cells that assume this function (scotopic vision). Both types of photoreceptors work coordinately in the interval between these values (mesopic vision). Each mechanism generates a different spectral sensibility. In this work, Citation: Navarrete-de Galvez, E.; the emission spectra of common sources in present public lighting installations are analyzed and Gago-Calderon, A.; Garcia-Ceballos, their normative photopic values translated to the corresponding mesopic condition, which more L.; Contreras-Lopez, M.A.; faithfully represents the vision mechanism of our eyes in these conditions. Based on a common Andres-Diaz, J.R. Adjustment of street urban configuration (ME6), we generated a large set of simulations to determine the ideal light Lighting Parameters from Photopic to point setup configuration (luminance and light point height vs. -

Birdshot Chorioretinopathy
Ocular Inflammation Service, Oxford Eye Hospital Birdshot Chorioretinopathy Information for patients What is birdshot chorioretinopathy? Birdshot chorioretinopathy (or retinochoroidopathy), normally shortened to ‘birdshot’, is a rare, potentially blinding, posterior uveitis. This is chronic inflammation of the choroid, which also tends to affect the retina and retinal vessels. It affects both eyes. In the picture below, you can see the position of the vitreous, retina and uvea (iris, ciliary body, pars planar and choroid), which has three sections; anterior, intermediate and posterior. The dotted area represents the uvea. Choroid Retina Vitreous Cornea Macula Lens Fovea Iris Optic nerve Ciliary body anterior intermediate posterior page 2 Birdshot chorioretinopathy is characterised by inflammation of the vitreous (clear jelly in the eye) which causes orange, yellow or cream coloured oval shaped spots at the back of your eye on your retina. These affect the macula (an area near the centre of the retina used for detailed vision) and can cause vision loss. The reason this disease is called ‘birdshot’ is because these spots look like the pattern seen when you fire birdshot pellets from a shotgun. What causes birdshot? It is believed to be due to an autoimmune disease. An autoimmune disease is an illness that occurs when the body tissues are attacked by its own immune system, which causes chronic inflammation. It is most likely to develop in people aged between 45 and 55, although it can also occur in much younger and older people. Birdshot is a relatively new disease. It was first discovered in 1949 and only given the title ‘birdshot’ in 1980. -

Vision Science Meets Visualization
Vision Science Meets Visualization Organizers: Christine Nothelfer Zoya Bylinskii Madison Elliott Cindy Xiong Danielle Albers Szafir Northwestern Massachusetts Institute University of British Northwestern University of Colorado University of Technology Columbia University Boulder Panelists: Ronald Rensink Todd Horowitz Steven Franconeri Karen Schloss Ruth Rosenholtz University of British National Cancer Northwestern University of Massachusetts Institute Columbia Institute University Wisconsin-Madison of Technology presenters each year. Recent developments in these research ABSTRACT topics (e.g., in visual attention, scene understanding, and quantity Vision science can explain what people see when looking at a perception) can inform our understanding of how people interpret visualization--what data features people attend to, what statistics visualized information. However, historically, few VIS they ascertain, and what they ultimately remember. These findings researchers have attended VSS, and few vision scientists have have significant relevance to visualization and can guide effective attended IEEE VIS. This limited overlap has stifled the exchange techniques and design practices. Intersections between of information, ideas, and questions between the two visualization and vision science have traditionally built upon communities. topics such as visual search, color perception, and pop-out. However, there is a broader space of vision science concepts that Further, while crossover between vision science and visualization could inform and explain ideas in visualization but no dedicated is not without precedent (e.g., work by Cleveland & McGill [2] venue for collaborative exchanges between the two communities. and Healey & Enns [8]), these interactions can benefit from This panel provides a space for this exchange by bringing five exposure to a broader set of vision science topics, such as object vision science experts to IEEE VIS to survey the modern vision tracking, ensemble statistics, and visual crowding. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Photopic and Scotopic – the “Eyes” Have It
Photopic and Scotopic – The “eyes” have it Dick Erdmann GE Specification Engineer In the human eye the perceived brightness of illumination depends of color. It takes more energy in the blue or red portion of the color spectrum to create the same sensation of brightness as in the yellow-green region. When it comes to sensing light in the human eye there are two main light-sensing cells called rods and cones. If one took a tube and looked straight ahead through it so that it allowed the field of view to be restricted 2 degrees, light photons would fall on the part of the eye called the fovea that consists mainly of cones. The peripheral area surrounding the fovea consists of both rods and cones with the rods outnumbering the cones about 10:1. Cones have a peak response in the yellow-green region of about 555 nano-meters and rods have a peak response in the bluish-green area of about 505 nano-meters. Because both the rods and cones have been shown to have different sensitivities to colors they can be represented by two different sensitivity curves called Photopic Curves (representing the cone) and Scotopic Curves (representing the rod). Years ago it was thought that the cones were responsible for daytime vision and rods for nighttime vision. Because of this, light meters that measure lighting levels such as footcandles, lumens, lux, etc. are weighted to the cone activated part of the eye ignoring the effect of rod-activated vision. But according to a study by Dr. Sam Berman and Dr. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment. -

Chromatic Function of the Cones D H Foster, University of Manchester, Manchester, UK
Chromatic Function of the Cones D H Foster, University of Manchester, Manchester, UK ã 2010 Elsevier Ltd. All rights reserved. Glossary length. If l is the path length and a(l) is the spectral absorptivity, then, for a homogeneous isotropic CIE, Commission Internationale de l’Eclairage – absorbing medium, A(l)=la(l) (Lambert’s law). The CIE is an independent, nonprofit organization Spectral absorptance a(l) – Ratio of the spectral responsible for the international coordination of radiant flux absorbed by a layer to the spectral lighting-related technical standards, including radiant flux entering the layer. If t(l) is the spectral colorimetry standards. transmittance, then a(l)=1Àt(l). The value of a(l) Color-matching functions – Functions of depends on the length or thickness of the layer. For a wavelength l that describe the amounts of three homogeneous isotropic absorbing medium, fixed primary lights which, when mixed, match a a(l)=1Àt(l)=1À10Àla(l), where l is the path length monochromatic light of wavelength l of constant and a(l) is the spectral absorptivity. Changes in the radiant power. The amounts may be negative. The concentration of a photopigment have the same color-matching functions obtained with any two effect as changes in path length. different sets of primaries are related by a linear Spectral absorptivity a(l) – Spectral absorbance of transformation. Particular sets of color-matching a layer of unit thickness. Absorptivity is a functions have been standardized by the CIE. characteristic of the medium, that is, the Fundamental spectral sensitivities – The color- photopigment. Its numerical value depends on the matching functions corresponding to the spectral unit of length.