1 Human Color Vision
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Solvent-Based Screen
Technical Data Sheet Nazdar 9600 Series Polyester Screen Ink v 12 EN 9600 Series Screen Ink is designed primarily for printing on untreated polyester films. The ink dries to a film exhibiting good gloss and flexibility with the exception of 9652 Super Opaque Ref: v 11 EN Black, which exhibits a matte, but flexible ink film. 9600 Series may be catalyzed with NB72 Catalyst or NB80 Adhesion Promoter for adhesion to a diverse range of substrates including polyester, some rubber, polycarbonate, melamine plastics, leather and some coated and uncoated metals. When printing 9600 Series on polycarbonate for insert‐mold decorating applications, up to 5% by weight addition of NB72 provides optimum performance. Substrates Maintain ink temperature at 65°‐90°F (18°‐32°C) for optimum print and cure performance. Lower Untreated polyester temperatures increase the ink viscosity, impairing Polyester coated surfaces flow and increasing film thickness. Elevated Some treated or top coated polyester temperatures lower the ink viscosity, reducing films print definition and film thickness. Polycarbonate Substrate recommendations are based on commonly available Pretest to determine optimum printing materials intended for the ink’s specific market when the inks parameters for a particular set of ink, substrate, are processed according to this technical data. While technical screen, press, and curing variables/conditions. information and advice on the use of this product is provided in good faith, the User bears sole responsibility for selecting Nazdar does not recommend inter‐mixing of 9600 Ink the appropriate product for their end‐use requirements. Series with other inks besides the 9600 Series. Reference the ‘Quality Statement’ at the end of this document. -

Conventional Screen Ink Color Chart
Conventional Screen Ink Color Chart 2700 Series, 5100 Series, 6100 Series, 7200 Series, 7700 Series, 7900 Series, 8400 Series, 8800 Series, 8900 Series, 9600 Series, 9700 Series, 9800 Series, ADE Series, PP Series, S2 Series. The Conventional Screen Inks Color Chart is representative of the colors available in several Nazdar Conventional ink series; however, some colors are not available in all ink series. Check the specific product pages for specific ink series color availability. PANTONE® Base Colors 2700 Series, 5100 Series, 6100 Series, 7200 Series, 7700 Series, 7900 Series, 8400 Series, 8800 Series, 8900 Series, 9600 Series, 9700 Series, 9800 Series, ADE Series, PP Series, S2 Series. conventiaonl The Conventional Screen Inks Color Chart is representative of the colors available in several Nazdar Conventional ink series; however, some colors are not available in all ink series. Check the specific product pages for specific ink series color availability. 60/360 61/361 62/362 63/363 64/364 65/365 Orange Yellow Warm Red Rubine Red Rhodamine Red Purple * PANTONE 165C * PANTONE 102C * PANTONE Warm Red C * PANTONE Rubine Red C * PANTONE Rhodamine Red C * PANTONE 2593C 66/366 67/367 68/368 69/369 Violet Reflex Blue Process Blue Green * PANTONE 2747C * PANTONE Reflex Blue C * PANTONE 3015 C * PANTONE Green C conventiaonl Color charts represented on Nazdar.com are provided to give printers a general idea of the colors available in Nazdar ink lines. Due to the wide variations and inconsistencies 10 in Primrose computer monitors and11 digitalLemon color palettes,12 the Medium colors seen here are 19only Fire approximations of20 actual Brilliant ink colors. -

Color Gamut of Halftone Reproduction*
Color Gamut of Halftone Reproduction* Stefan Gustavson†‡ Department of Electrical Engineering, Linkøping University, S-581 83 Linkøping, Sweden Abstract tern then gets attenuated once more by the pattern of ink that resides on the surface, and the finally reflected light Color mixing by a halftoning process, as used for color is the result of these three effects combined: transmis- reproduction in graphic arts and most forms of digital sion through the ink film, diffused reflection from the hardcopy, is neither additive nor subtractive. Halftone substrate, and transmission through the ink film again. color reproduction with a given set of primary colors is The left-hand side of Fig. 2 shows an exploded view of heavily influenced not only by the colorimetric proper- the ink layer and the substrate, with the diffused reflected ties of the full-tone primaries, but also by effects such pattern shown on the substrate. The final viewed image as optical and physical dot gain and the halftone geom- is a view from the top of these two layers, as shown to etry. We demonstrate that such effects not only distort the right in Fig. 2. The dots do not really increase in the transfer characteristics of the process, but also have size, but they have a shadow around the edge that makes an impact on the size of the color gamut. In particular, a them appear larger, and the image is darker than what large dot gain, which is commonly regarded as an un- would have been the case without optical dot gain. wanted distortion, expands the color gamut quite con- siderably. -

Working with Halftones
Making Halftone negatives First convert your document to a bitmap file. When printing on an Epson choose 1440 dpi. When printing to a laser printer choose 1200 dpi. When you get the Option window choose Method -> Halftone Screen. Now choose your halftone screen frequency, which may be around 100 lpi for commercial printing, or for an art project. To reduce the likelihood of a moire pattern, it is best to specify different screen angles for each color. Traditionally these were set at 45º for Cyan, 75º for Magenta, 90º for Yellow and 105º for Black. Notice the darker colors are 30º from each other and yellow is set halfway between Magenta and Black. If you’re outputting a duo or tri tone set each 30º from each other. For dot shape, the ellipse halftone dot is often recommended for screen-printing. The following are from http://www.kevinhaas.com Creating a Halftone Bitmap 1. You will need a grayscale file that is at least 150ppi at the size you will print it. This information can be checked by going to Image > Image Size... in Photoshop. 2. Convert the image into a Bitmap: Go to Image > Mode > Bitmap. Set your Output Resolution to 720 and Method to ‘Halftone’. The Output Resolution should be your lpi x 16, and evenly divisible by your printer’s maximum output resolution. For example, a 35lpi halftone needs an Output Resolution of at least 560, but 720 is the next higher resolution that is the inkjet printer’s resolution of 1440 evenly divided by 2. 3. -

Federal Wage System Glossary of Printing Terms for the 4400 Job Family
Glossary of Printing Terms for the 4400 Job Family TS-45 October 1981 Federal Wage System Glossary of Printing Terms for the 4400 Job Family The terms and definitions contained in this glossary are intended to facilitate the application of published job grading standards to occupations in the Printing Family. The glossary does not represent a comprehensive listing of technical terms and processes common to printing occupations. Additional information may be found in dictionaries, and technical publications such as agency guides, trade magazines, and technical manuals. Aluminum Plate. A thin sheet of aluminum used in lithography for some press plates; image applied photographically; used for both surface-type and deep-etch offset plates. Aperture. A small opening in a plate or sheet. In cameras, the aperture is usually variable in the form of an iris diaphragm and regulates the amount of light which passes through the lens. The working aperture is the diameter of that part of the lens actually used. Asphaltum. A bituminous mixture used as an acid resist or protectant in photomechanics. In lithography, used to make printing image on press plate permanently ink-receptive. Autoscreen (Film). A photographic film embodying the halftone screen; exposed to a continuous-tone image, produces a dot pattern automatically just as if a halftone screen had been used in the camera. Backing-Up. Printing the other side, of a printed sheet. Back Pressure. The squeeze pressure between the blanket (offset) cylinder and the impression cylinder; sometimes called "impression pressure." Base Color. A first color used as a background on which other colors are printed. -

Applications of High Dynamic Range Imaging Stereo Matching and Mesopic Rendering
Applications of High Dynamic Range Imaging Stereo Matching and Mesopic Rendering PhD THESIS submitted in partial fulfillment of the requirements for the degree of Doctor of Technical Sciences within the Vienna PhD School of Informatics by Eng. Tara Akhavan Registration Number 1128895 to the Faculty of Informatics at the Vienna University of Technology Advisor: Assoc. Prof. Mag. Dr. Hannes Kaufmann External reviewers: . Alan Chalmers, WMG, University of Warwick, United Kingdom. Kadi Bouatouch, ISTIC, University of Rennes I, France. Vienna, 8th April, 2017 Tara Akhavan Hannes Kaufmann Technische Universität Wien A-1040 Wien Karlsplatz 13 Tel. +43-1-58801-0 www.tuwien.ac.at Declaration of Authorship Eng. Tara Akhavan Address I hereby declare that I have written this Doctoral Thesis independently, that I have completely specified the utilized sources and resources and that I have definitely marked all parts of the work - including tables, maps and figures - which belong to other works or to the internet, literally or extracted, by referencing the source as borrowed. Vienna, 8th April, 2017 Tara Akhavan iii Acknowledgements Firstly, I would like to express my deepest gratitude to my supervisor Hannes Kaufmann for his unlimited support, inspiration, motivation, belief, and guidance. My special thanks to Christian Breiteneder who was always there to help resolving hardest problems with wisdom and vision. I am very thankful to Alan Chalmers and Kadi Bouatouch who agreed to be my thesis external reviewers but most importantly planned valuable workshops through the course of my PhD and invited me to network with and learn from the experts in the field. Part of my PhD research was conducted at Tandemlaunch Inc. -
Chapter 6 COLOR and COLOR VISION
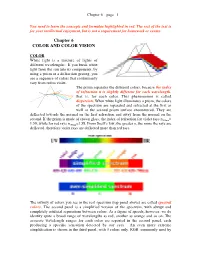
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

A Guide to Colorimetry
A Guide to Colorimetry Sherwood Scientific Ltd 1 The Paddocks Cherry Hinton Road Cambridge CB1 8DH England www.sherwood-scientific.com Tel: +44 (0)1223 243444 Fax: +44 (0)1223 243300 Registered in England and Wales Registration Number 2329039 Reg. Office as above Group I II III IV V VI VII VIII Period 1A 8A 1 2 1 H 2A 3A 4A 5A 6A 7A He 1.008 4.003 3 4 5 6 7 8 9 10 2 Li Be B C N O F Ne 6.939 9.0122 10.811 12.011 14.007 15.999 18.998 20.183 11 12 13 14 15 16 17 18 3 Na Mg 3B 4B 5B 6B 7B [---------------8B-------------] 1B 2B Al Si P S Cl Ar 22.99 24.312 26.982 28.086 30.974 32.064 35.453 39.948 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 4 K Ca Sc Ti V Cr Mn Fe Co Ni Cu Zn Ga Ge As Se Br Kr 39.102 40.08 44.956 47.9 50.942 51.996 54.938 55.847 58.933 58.71 63.546 65.37 69.72 72.59 74.922 78.96 79.904 83.8 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 5 Rb Sr Y Zr Nb Mo Tc Ru Rh Pd Ag Cd In Sn Sb Te I Xe 85.47 87.52 88.905 91.22 92.906 95.94 [97] 101.07 102.91 106.4 107.87 112.4 114.82 118.69 121.75 127.6 126.9 131.3 55 56 57* 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 6 Cs Ba La Hf Ta W Re Os Ir Pt Au Hg Ti Pb Bi Po At Rn 132.91 137.34 138.91 178.49 180.95 183.85 186.2 190.2 192.2 195.09 196.97 200.59 204.37 207.19 208.98 209 210 222 87 88 89** 104 105 106 107 108 109 110 111 112 114 116 7 Fr Ra Ac Rt Db Sg Bh Hs Mt 215 226.03 227.03 [261] [262] [266] [264] [269] [268] [271] [272] [277] [289] [289] 58 59 60 61 62 63 64 65 66 67 68 69 70 71 * Lanthanides Ce Pr Nd Pm Sm Eu Gd Tb Dy Ho Er Tm Yb Lu 140.12 140.91 144.24 145 150.35 151.96 157.25 158.92 152.5 164.93 167.26 168.93 173.04 174.97 90 91 92 93 94 95 96 97 98 99 100 101 102 103 ** Actinides Th Pa U Np Pu Am Cm Bk Cf Es Fm Md No Lr 232.04 231 238 237.05 239.05 241.06 244.06 249.08 251 252.08 257.1 258.1 259.1 262.11 The ability to analyse and quantify colour in aqueous solutions and liquids using a colorimeter is something today’s analyst takes for granted. -

Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs
sustainability Article Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs Enrique Navarrete-de Galvez 1 , Alfonso Gago-Calderon 1,* , Luz Garcia-Ceballos 2, Miguel Angel Contreras-Lopez 2 and Jose Ramon Andres-Diaz 1 1 Proyectos de Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (E.N.-d.G.); [email protected] (J.R.A.-D.) 2 Expresión Gráfica en la Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (L.G.-C.); [email protected] (M.A.C.-L.) * Correspondence: [email protected]; Tel.: +34-951-952-268 Abstract: The sensitivity of the human eye varies with the different lighting conditions to which it is exposed. The cone photoreceptors perceive the color and work for illuminance conditions greater than 3.00 cd/m2 (photopic vision). Below 0.01 cd/m2, the rods are the cells that assume this function (scotopic vision). Both types of photoreceptors work coordinately in the interval between these values (mesopic vision). Each mechanism generates a different spectral sensibility. In this work, Citation: Navarrete-de Galvez, E.; the emission spectra of common sources in present public lighting installations are analyzed and Gago-Calderon, A.; Garcia-Ceballos, their normative photopic values translated to the corresponding mesopic condition, which more L.; Contreras-Lopez, M.A.; faithfully represents the vision mechanism of our eyes in these conditions. Based on a common Andres-Diaz, J.R. Adjustment of street urban configuration (ME6), we generated a large set of simulations to determine the ideal light Lighting Parameters from Photopic to point setup configuration (luminance and light point height vs. -

Vision Science Meets Visualization
Vision Science Meets Visualization Organizers: Christine Nothelfer Zoya Bylinskii Madison Elliott Cindy Xiong Danielle Albers Szafir Northwestern Massachusetts Institute University of British Northwestern University of Colorado University of Technology Columbia University Boulder Panelists: Ronald Rensink Todd Horowitz Steven Franconeri Karen Schloss Ruth Rosenholtz University of British National Cancer Northwestern University of Massachusetts Institute Columbia Institute University Wisconsin-Madison of Technology presenters each year. Recent developments in these research ABSTRACT topics (e.g., in visual attention, scene understanding, and quantity Vision science can explain what people see when looking at a perception) can inform our understanding of how people interpret visualization--what data features people attend to, what statistics visualized information. However, historically, few VIS they ascertain, and what they ultimately remember. These findings researchers have attended VSS, and few vision scientists have have significant relevance to visualization and can guide effective attended IEEE VIS. This limited overlap has stifled the exchange techniques and design practices. Intersections between of information, ideas, and questions between the two visualization and vision science have traditionally built upon communities. topics such as visual search, color perception, and pop-out. However, there is a broader space of vision science concepts that Further, while crossover between vision science and visualization could inform and explain ideas in visualization but no dedicated is not without precedent (e.g., work by Cleveland & McGill [2] venue for collaborative exchanges between the two communities. and Healey & Enns [8]), these interactions can benefit from This panel provides a space for this exchange by bringing five exposure to a broader set of vision science topics, such as object vision science experts to IEEE VIS to survey the modern vision tracking, ensemble statistics, and visual crowding. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

S Selection of Intraocular Lens on the Basis of Light Transmittance Properties
Eye (2017) 31, 258–272 OPEN Official journal of The Royal College of Ophthalmologists www.nature.com/eye 1 2 2 2 REVIEW The evidence informing XLi, D Kelly , JM Nolan , JL Dennison and S Beatty2,3 the surgeon’s selection of intraocular lens on the basis of light transmittance properties Abstract In recent years, manufacturers and distributors surgical procedure worldwide,1 and the need have promoted commercially available intrao- for such surgery is likely to rise because cular lenses (IOLs) with transmittance proper- of increasing longevity.2 Commercially fi ties that lter visible short-wavelength (blue) available IOLs can vary in terms of material light on the basis of a putative photoprotective (polymethylmethacrylate, polymers, silicone effect. Systematic literature review. Out of or acrylic),3,4 hydrophobicity (hydrophobic vs 21 studies reporting on outcomes following hydrophilic),5 sphericity (aspheric vs spheric)6 implantation of blue-light-filtering IOLs (invol- 1 and light-filtering properties. In this paper, Pharmaceutical & ving 8914 patients and 12 919 study eyes Molecular Biotechnology undergoing cataract surgery), the primary out- we review the evidence germane to the Research Centre, fi come was vision, sleep pattern, and photopro- putative and relative risks and/or bene ts of Department of Chemical & fi Life Sciences, Waterford tection in 9 (42.9%), 9 (42.9%), and 3 (14.2%) implanting IOLs that lter visible short- Institute of Technology, respectively, and, of these, only 7 (33.3%) can be wavelength light (ie, o500 nm), referred to as Waterford, Ireland classed as high as level 2b (individual cohort blue-light-filtering IOLs for the purpose of this study/low-quality randomized controlled trials), review.