Predicting the Invasion Potential of Indigenous Restricted Mango Fruit Borer, Citripestis Eutraphera (Lepidoptera: Pyralidae) in India Based on Maxent Modelling
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Insert Report Title Here
Vegetable biosecurity & quarantine gap analysis Prue McMichael Scholefield Robinson Horticultural Services Pty Ltd Project Number: VG07087 VG07087 This report is published by Horticulture Australia Ltd to pass on information concerning horticultural research and development undertaken for the vegetable industry. The research contained in this report was funded by Horticulture Australia Ltd with the financial support of the vegetable industry. All expressions of opinion are not to be regarded as expressing the opinion of Horticulture Australia Ltd or any authority of the Australian Government. The Company and the Australian Government accept no responsibility for any of the opinions or the accuracy of the information contained in this report and readers should rely upon their own enquiries in making decisions concerning their own interests. ISBN 0 7341 1849 X Published and distributed by: Horticulture Australia Ltd Level 7 179 Elizabeth Street Sydney NSW 2000 Telephone: (02) 8295 2300 Fax: (02) 8295 2399 E-Mail: [email protected] © Copyright 2008 FINAL REPORT Vegetable Biosecurity and Quarantine Gap Analysis VG07087 Prepared for : Horticulture Australia Ltd HAL Project No. VG07087 Prepared by : Prue McMichael Completion Date : September 2008 SCHOLEFIELD ROBINSON HORTICULTURAL SERVICES PTY LTD 118A Glen Osmond Road, Parkside SA 5063 Australia ACN 008 199 737 PO Box 650, Fullarton SA 5063 Ph: (08) 8373 2488 ABN 63 008 199 737 Fax: (08) 8373 2442 Email: [email protected] Web Site: www.srhs.com.au Offices in Adelaide and Mildura Scholefield Robinson Horticultural Services Pty Ltd HAL Project No. VG 07087 PROJECT LEADER Dr Prue McMichael Senior Consultant/Plant Pathologist Scholefield Robinson Horticultural Services Pty Ltd PO Box 650 Fullarton SA 5063 PURPOSE OF REPORT This Final Report has been prepared to document information acquired, analysed and considered during the review undertaken for HAL, into all aspects of the biosecurity of Australia’s vegetable industries that are members of AUSVEG. -

Citrus Industry Biosecurity Plan 2015
Industry Biosecurity Plan for the Citrus Industry Version 3.0 July 2015 PLANT HEALTH AUSTRALIA | Citrus Industry Biosecurity Plan 2015 Location: Level 1 1 Phipps Close DEAKIN ACT 2600 Phone: +61 2 6215 7700 Fax: +61 2 6260 4321 E-mail: [email protected] Visit our web site: www.planthealthaustralia.com.au An electronic copy of this plan is available through the email address listed above. © Plant Health Australia Limited 2004 Copyright in this publication is owned by Plant Health Australia Limited, except when content has been provided by other contributors, in which case copyright may be owned by another person. With the exception of any material protected by a trade mark, this publication is licensed under a Creative Commons Attribution-No Derivs 3.0 Australia licence. Any use of this publication, other than as authorised under this licence or copyright law, is prohibited. http://creativecommons.org/licenses/by-nd/3.0/ - This details the relevant licence conditions, including the full legal code. This licence allows for redistribution, commercial and non-commercial, as long as it is passed along unchanged and in whole, with credit to Plant Health Australia (as below). In referencing this document, the preferred citation is: Plant Health Australia Ltd (2004) Industry Biosecurity Plan for the Citrus Industry (Version 3.0 – July 2015). Plant Health Australia, Canberra, ACT. Disclaimer: The material contained in this publication is produced for general information only. It is not intended as professional advice on any particular matter. No person should act or fail to act on the basis of any material contained in this publication without first obtaining specific and independent professional advice. -
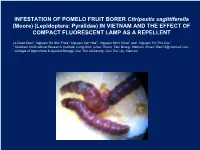
INFESTATION of POMELO FRUIT BORER Citripestis Sagittiferella (Moore) (Lepidoptera: Pyralidae) in VIETNAM and the EFFECT of COMPACT FLUORESCENT LAMP AS a REPELLENT
INFESTATION OF POMELO FRUIT BORER Citripestis sagittiferella (Moore) (Lepidoptera: Pyralidae) IN VIETNAM AND THE EFFECT OF COMPACT FLUORESCENT LAMP AS A REPELLENT Le Quoc Dien1, Nguyen Thi Kim Thoa1, Nguyen Van Hoa1 , Nguyen Minh Chau1 and Nguyen Thi Thu Cuc2 1 Southern Horticultural Research Institute, Long Dinh, Chau Thanh, Tien Giang, Vietnam, Email: Dien72@ hotmail.com. 2 College of Agriculture & Applied Biology, Can Tho University, Can Tho city, Vietnam 1. INTRODUCTION Pomelo culture has significantly played a key role in the Mekong Delta economy The Mekong Delta accounted for about 85% of the Viet Nam citrus production (MARD, 2010). 2011, pomelo fruit borer Citripestis sagittiferella (Moore) has started spreading and causing severe damage across all planted areas in Mekong delta as Ben Tre, Tien Giang, Soc Trang, Vinh Long, Bac Lieu, Can Thơ city and spacial và specially on Hau Giang (PPD, 2013). This specices also has started spreading Indonesia, Philippines, Thai Land and Malaysia. Control fruit borer ? Insect parasite, Rhoptromeris sp. (Hymenoptera: Eucoilidae) plays an important role in the control of Citripestis sagittiferrella on lemon at University Pertanian Malaysia orchard. At 24% larval parasitism was estimated from larvae collelcted in the field. Field studies to evaluate the effectiveness of malathion 50 EC emulsifiable concentrate sprayed to trees at 0.1% a.i. active ingredient show that the percentage of fruits attacked was nuch more higher in the sprayed block than that of the unsprayed block (Hussein and Rahman (1981). Insect visual systems to find out about the abililty to detect objects, colours, movement, prey or conspecifics using the properties of light provides an incredible spatial advantage as critical information is collected and responded to from an extended scene. -

Field Guide to Pests, Beneficials, Diseases and Disorders of Mangoes
E D I U G D L E I F Pests, Beneficials, Diseases and Disorders of Mangoes PDF View, search and navigate This PDF has been created specifically for on-screen display, e-mail, and internet use. SCREEN VIEW The page view of this electronic document has been automatically set to ‘Fit Screen Page’ and viewed as facing pages. To view as single pages go to: VIEW tab > Select PAGE LAYOUT > Choose Single page. Viewing options can be modified using the shortcut keys below. SHORTCUT KEYS ACTION Ctrl+0 (Zero) FIT PAGE Ctrl+M Zooms to selected sizes Ctrl+L FULL SCREEN MODE SEARCH OPTIONS Ctrl+F Search for keywords or terms Shortcuts provided may vary depending on software installed on your computer. NAVIGATION BOOKMARKS - The Bookmark panel located on the left hand side can be used to jump to topic destinations in the PDF (just click on bookmark listing). QUICK LINKS - Topics listed on the contents page have quick links and when clicked on will take you directly to the topic page. PRINT OPTIONS For optimal print it is recommended that this document is printed double sided on A4 paper (select portrait and flip on long edge settings), set your print page scaling to ‘Fit to Printer Margins'. Ctrl+P To Print Choose File > Print Setup (Windows) or File > Page Setup (Mac OS) choose A4 paper size, page orientation (portrait), and other general printing options including instructions mentioned above. Print options will vary on diffferent printers and drivers being used. FIELD GUIDE to Pests, Beneficials, Diseases and Disorders of Mangoes December 2010 Northern Territory Government Department of Resources GPO Box 3000, Darwin NT 0801, AUSTRALIA © Copyright, Northern Territory Government 2010 Disclaimer While all care has been taken to ensure that information contained in this booklet is true and correct at the time of publication, the Northern Territory of Australia gives no warranty or assurance, and makes no representation as to the accuracy of any information or advice contained in this publication, or that it is suitable for your intended use. -

EU Project Number 613678
EU project number 613678 Strategies to develop effective, innovative and practical approaches to protect major European fruit crops from pests and pathogens Work package 1. Pathways of introduction of fruit pests and pathogens Deliverable 1.3. PART 7 - REPORT on Oranges and Mandarins – Fruit pathway and Alert List Partners involved: EPPO (Grousset F, Petter F, Suffert M) and JKI (Steffen K, Wilstermann A, Schrader G). This document should be cited as ‘Grousset F, Wistermann A, Steffen K, Petter F, Schrader G, Suffert M (2016) DROPSA Deliverable 1.3 Report for Oranges and Mandarins – Fruit pathway and Alert List’. An Excel file containing supporting information is available at https://upload.eppo.int/download/112o3f5b0c014 DROPSA is funded by the European Union’s Seventh Framework Programme for research, technological development and demonstration (grant agreement no. 613678). www.dropsaproject.eu [email protected] DROPSA DELIVERABLE REPORT on ORANGES AND MANDARINS – Fruit pathway and Alert List 1. Introduction ............................................................................................................................................... 2 1.1 Background on oranges and mandarins ..................................................................................................... 2 1.2 Data on production and trade of orange and mandarin fruit ........................................................................ 5 1.3 Characteristics of the pathway ‘orange and mandarin fruit’ ....................................................................... -

1. Padil Species Factsheet Scientific Name: Common Name Image
1. PaDIL Species Factsheet Scientific Name: Citripestis eutraphera (Meyrick) 1933 (Lepidoptera:Pyralidae) Common Name Mango Fruit Borer Live link: http://www.padil.gov.au/pests-and-diseases/Pest/Main/142262 Image Library Australian Biosecurity Live link: http://www.padil.gov.au/pests-and-diseases/ Partners for Australian Biosecurity image library Department of Agriculture, Water and the Environment https://www.awe.gov.au/ Department of Primary Industries and Regional Development, Western Australia https://dpird.wa.gov.au/ Plant Health Australia https://www.planthealthaustralia.com.au/ Museums Victoria https://museumsvictoria.com.au/ 2. Species Information 2.1. Details Specimen Contact: daff.gov.au, nt.gov.au - daff.gov.au, nt.gov.au Author: S. Anderson & L. Tran-Nguyen Citation: S. Anderson & L. Tran-Nguyen (2012) Mango Fruit Borer(Citripestis eutraphera)Updated on 2/24/2012 Available online: PaDIL - http://www.padil.gov.au Image Use: Free for use under the Creative Commons Attribution-NonCommercial 4.0 International (CC BY- NC 4.0) 2.2. URL Live link: http://www.padil.gov.au/pests-and-diseases/Pest/Main/142262 2.3. Facets Commodity Overview: Field Crops and Pastures Commodity Type: Mango Distribution: Australasian - Oceanian, South and South-East Asia Group: Moths Status: Exotic Species Establishment in Australia 2.4. Other Names Citripestis eutraphera (Meyrick 1933:388) (Myelois) Mango Fruit Borer Philotroctis eutraphera Meyrick, 1933 by monotypy Roesler 1983:19 2.5. Diagnostic Notes **Identification is currently undergoing peer review as part of the National Diagnostic Protocols by SPHDS.** Characteristic damage includes spoiled fruit, holes in the side of fruit with frass and sap; premature fruit drop can occur as the larvae approach maturity (L Zhang 2010 pers. -

The Major Arthropod Pests and Weeds of Agriculture in Southeast Asia
The Major Arthropod Pests and Weeds of Agriculture in Southeast Asia: Distribution, Importance and Origin D.F. Waterhouse (ACIAR Consultant in Plant Protection) ACIAR (Australian Centre for International Agricultural Research) Canberra AUSTRALIA The Australian Centre for International Agricultural Research (ACIAR) was established in June 1982 by an Act of the Australian Parliament. Its mandate is to help identify agricultural problems in developing countries and to commission collaborative research between Australian and developing country researchers in fields where Australia has a special research competence. Where trade names are used this constitutes neither endorsement of nor discrimination against any product by the Centre. ACIAR MO'lOGRAPH SERIES This peer-reviewed series contains the results of original research supported by ACIAR, or deemed relevant to ACIAR's research objectives. The series is distributed internationally, with an emphasis on the Third World. © Australian Centre for 1I1lernational Agricultural Resl GPO Box 1571, Canberra, ACT, 2601 Waterhouse, D.F. 1993. The Major Arthropod Pests an Importance and Origin. Monograph No. 21, vi + 141pI- ISBN 1 86320077 0 Typeset by: Ms A. Ankers Publication Services Unit CSIRO Division of Entomology Canberra ACT Printed by Brown Prior Anderson, 5 Evans Street, Burwood, Victoria 3125 ii Contents Foreword v 1. Abstract 2. Introduction 3 3. Contributors 5 4. Results 9 Tables 1. Major arthropod pests in Southeast Asia 10 2. The distribution and importance of major arthropod pests in Southeast Asia 27 3. The distribution and importance of the most important arthropod pests in Southeast Asia 40 4. Aggregated ratings for the most important arthropod pests 45 5. Origin of the arthropod pests scoring 5 + (or more) or, at least +++ in one country or ++ in two countries 49 6. -

Lepidoptera : Pyralidae
REVIEW ZOOS' PRINT JOURNAL 21(5): 2245-2258 AN INVENTORY OF INDIAN PYRALIDS (LEPIDOPTERA: PYRALIDAE) George Mathew Division of Entomology, Kerala Forest Research Institute, Peechi, Kerala 680653, India Email: [email protected] ABSTRACT combination has been given. Altogether, 1646 species (against An inventory of 1646 species of pyralids so far recorded 1136 species reported in the Fauna of British India) is given. from India is presented indicating the year of publication While every attempt has been made to cover as many species and the current nomenclatural combination. A key adapted from literature is also given for separation of subfamilies. as possible, it is possible that some names could have been left out due to oversight. It is hoped that the present list may serve KEYWORDS as a draft for immediate reference until a detailed revision on Checklist, India, inventory, Lepidoptera, moths, Pyralidae, this group is prepared. subfamily key The classification followed in this work is the one proposed by Due to discovery of new species and also due to refinements in Munroe (1972) for the Pyralidae of America north of Mexico. the taxonomic techniques, considerable changes have taken Munroe and Solis (1999) have also given a detailed taxonomic place in the taxonomic status of various categories of the Indian treatment of this group. A key adapted from the above work is Pyralidae since publication of Sir George Hampson’s Fauna presented here for the separation of various subfamilies. volume in 1896 in the ‘Fauna of British India’ series. As a result, the nomenclature of a number of species had undergone REFERENCES changes, some times on several occasions and unaware of these Arora, G.S. -
Species Composition of Herbivorous Insects and Ants on Trees in The
Sci. Rep. Kanazawa Univ. ARTICLES Vol. 63, pp. 45-58, 2019 Species composition of herbivorous insects and ants on trees in the plantations of durian Durio zibethinus and citrus fruits Citrus amblycarpa in Tarakan Island of Borneo 1, 2 * 1 Abdul RAHIM and Kyohsuke OHKAWARA 1 Laboratory of Ecology, Division of Biological Sciences, Graduate School of Natural Science and Technology, Kanazawa University, Kanazawa 920-1192, Japan 2 Department of Agrotechnology, Agriculture Faculty, Borneo University, Tarakan, North Kalimantan 77123, Indonesia (Received January 24, 2019 and accepted in revised form February 26, 2019) Abstract In the tropics, the specific insect communities are observed in the fruit plantations as one form of agroforestry systems. The information on the communities of herbivorous insects as pests and ants as pest-control agents could be useful in facilitating effective management of plantations. Durian Durio zibethinus and citrus fruits Citrus amblycarpa are traditionally grown as agricultural products in the North Kalimantan region of Borneo, though the information on herbivorous insects and ants remains scant. In this study, we assessed the species composition and densities of herbivorous insects and ants inhabiting on trees in the plantations of durian and citrus fruits in Tarakan Island of North Kalimantan. From 2016 to 2017, 55649 individuals of herbivores comprised of 61 species and 64360 ant workers comprised of 25 species were collected by tree- beating sampling on 59 durian and 63 citrus trees in three plantation sites (A-C). In citrus fruit trees, aphid Toxoptera citridida was dominant in the herbivore communities. While, in durian trees, more than 80% of herbivores was occupied by various herbivore groups, leaf beetle, moth, mealybug, psyllid and leafhopper. -
PRA Report of Mango, SPCBP, PQW, DAE.Pdf
Government of the People’s Republic of Bangladesh Office of the Project Director Strengthening Phytosanitary Capacity in Bangladesh Project Plant Quarantine Wing Department of Agriculture Extension Khamarbari, Farmgate, Dhaka-1205 REPORT ON PEST RISK ANALYSIS (PRA) OF MANGO IN BANGLADESH DEVELOPMENT TECHNICAL CONSULTANTS PVT. LTD. (DTCL) Gulshan-1, Dhaka, Bangladesh DTCL Government of the People’s Republic of Bangladesh Ministry of Agriculture Office of the Project Director Strengthening Phytosanitary Capacity in Bangladesh Project Plant Quarantine Wing Department of Agriculture Extension Khamarbari, Farmgate, Dhaka-1205 REPORT ON PEST RISK ANALYSIS (PRA) OF MANGO IN BANGLADESH Development Technical Consultants Pvt. Ltd. (DTCL) DTCL JB House, Plot-62, Road-14/1, Block-G, Niketon Gulshan -1, Dhaka-1212, Bangladesh Tel: +880 2 9856438, 9856439 Fax: +880 2 9840973 E-mail: [email protected] Website: www.dtcltd.com MAY 2015 TEAM OF CONSULTANTS CONTRIBUTORS TO PEST RISK ANALYSIS OF MANGO IN BANGLADESH Team Members Position in the Team 1. Prof. Dr. Md. Razzab Ali Team Leader Department of Entomology Sher-e-Bangla Agricultural University Dhaka-1207, Bangladesh 2. Prof. Dr. Md. Ramiz Uddin Miah Entomologist Department of Entomology Bangabandhu Sheikh Mujibur Rahman Agricultural University Gazipur, Bangladesh 3. Prof. Dr. M. Salah Uddin M. Chowdhury Plant Pathologist Department of Plant Pathology Sher-e-Bangla Agricultural University Dhaka-1207, Bangladesh 4. Prof. Dr. Md. Abdul Karim Agronomist Department of Agronomy Bangabandhu Sheikh Mujibur Rahman Agricultural University Gazipur, Bangladesh 5. Dr. B. A. A. Mustafi Agriculture Economist Ex-Director (Research) Bangladesh Rice Research Institute Gazipur, Bangladesh 6. Dr. M. M. Amir Hossain Backstopping Consultant Managing Director Development Technical Consultants Pvt. -

3410-34-P DEPARTMENT of AGRICULTURE Animal and Plant
This document is scheduled to be published in the Federal Register on 03/29/2018 and available online at https://federalregister.gov/d/2018-06288, and on FDsys.gov BILLING CODE: 3410-34-P DEPARTMENT OF AGRICULTURE Animal and Plant Health Inspection Service 7 CFR Part 319 [Docket No. APHIS-2016-0034] RIN 0579-AE33 Importation of Pummelo From Thailand Into the Continental United States AGENCY: Animal and Plant Health Inspection Service, USDA. ACTION: Proposed rule. SUMMARY: We are proposing to amend the regulations to allow the importation of fresh pummelo fruit from Thailand into the continental United States. As a condition of entry, fresh pummelo fruit from Thailand would be subject to a systems approach that would include irradiation treatment, packinghouse processing requirements, and port of entry inspection. The fruit would also be required to be imported in commercial consignments and be accompanied by a phytosanitary certificate issued by the national plant protection organization of Thailand. This action would allow for the importation of fresh pummelo fruit from Thailand while continuing to provide protection against the introduction of plant pests into the continental United States. DATES: We will consider all comments that we receive on or before [Insert date 60 days after date of publication in the Federal Register]. ADDRESSES: You may submit comments by either of the following methods: Federal eRulemaking Portal: Go to http://www.regulations.gov/#!docketDetail;D=APHIS-2016-0034. Postal Mail/Commercial Delivery: Send your comment to Docket No. APHIS-2016- 0034, Regulatory Analysis and Development, PPD, APHIS, Station 3A-03.8, 4700 River Road Unit 118, Riverdale, MD 20737-1238. -

Scientific Panel on Plant Health
Animal Health and Plant Health Unit Scientific Panel on Plant Health MINUTES OF THE 81st MEETING OF THE WORKING GROUP ON ARTHROPOD PEST CATEGORISATION WEB-conference 8th of September 2021 (Agreed on 9th of September 2021) Participants Working Group Members: Alan MacLeod (Chair) Josep Anton Jaques Miret (vice-Chair) Lucia Zappalà (vice-Chair) Jean-Claude Grégoire (Université Libre de Bruxelles (ULB)) Hearing Experts1: Not Applicable European Commission and/or Member States representatives: Not Applicable EFSA: Animal and Plant Health Unit: Ewelina Czwienczek, Virag Kertesz 1. Welcome and apologies for absence Apologies were received from Chris Malumphy (FERA) 2. Adoption of agenda The agenda was adopted without changes. 3. Declarations of Interest of Working Groups members 1 As defined in Article 17 of the Decision of the Executive Director concerning the selection of members of the Scientific Committee, the Scientific Panels, and the selection of external experts to assist EFSA with its scientific work: http://www.efsa.europa.eu/en/keydocs/docs/expertselection.pdf. European Food Safety Authority Via Carlo Magno 1A – 43126 Parma, Italy Tel. +39 0521 036 111 │ www.efsa.europa.eu In accordance with EFSA’s Policy on Independence2 and the Decision of the Executive Director on Competing Interest Management3, EFSA screened the Annual Declarations of Interest filled out by the Working Group members invited to the present meeting. No Conflicts of Interest related to the issues discussed in this meeting have been identified during the screening process, and no interests were declared orally by the members at the beginning of this meeting. 4. Agreement of the minutes of the 80th Working Group meeting held on 16thof July2021.