Full Text (3628.4
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Silurian Rugose Corals of the Central and Southwest Great Basin
Silurian Rugose Corals of the Central and Southwest Great Basin GEOLOGICAL SURVEY PROFESSIONAL PAPER 777 Silurian Rugose Corals of the Central and Southwest Great Basin By CHARLES W. MERRIAM GEOLOGICAL SURVEY PROFESSIONAL PAPER 777 A stratigraphic-paleontologic investigation of rugose corals as aids in age detern2ination and correlation of Silurian rocks of the Great Basin with those of other regions UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1973 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress catalog-card No. 73-600082 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402- Price $2.15 (paper cover) Stock Number 2401-00363 CONTENTS Page Page Abstract--------------------------------------------------------------------------- 1 Systematic and descriptive palaeontology-Continued Introduction -------------------------------------------------------------------- 1 Family Streptelasmatidae-Continued Purpose and scope of investigation------------------------------- 1 Dalmanophyllum ------------------------------------------------- 32 History of investigation ---------------------------------------------- 1 Family Stauriidae ------------------------------------------------------- 32 Methods of study------------------------------------------------------- 2 Cyathoph y llo ides-------------------------------------------------- 32 Acknowledgments------------------------------------------------------- 4 Palaeophyllum -
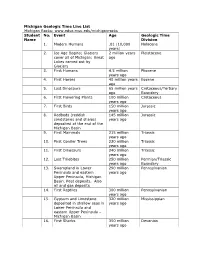
Michigan Geologic Time Line List Michigan Rocks: Student No
Michigan Geologic Time Line List Michigan Rocks: www.educ.msu.edu/michiganrocks Student No. Event Age Geologic Time Name Division 1. Modern Humans .01 (10,000 Holocene years) 2. Ice Age Begins; Glaciers 2 million years Pleistocene cover all of Michigan; Great ago Lakes carved out by Glaciers 3. First Humans 4.5 million Pliocene years ago 4. First Horses 40 million years Eocene ago 5. Last Dinosaurs 65 million years Cretaceous/Tertiary ago Boundary 6. First Flowering Plants 100 million Cretaceous years ago 7. First Birds 150 million Jurassic years ago 8. Redbeds (reddish 145 million Jurassic sandstones and shales) years ago deposited at the end of the Michigan Basin 9. First Mammals 215 million Triassic years ago 10. First Conifer Trees 230 million Triassic years ago 11. First Dinosaurs 240 million Triassic years ago 12. Last Trilobites 250 million Permian/Triassic years ago Boundary 13. Swampland in Lower 290 million Pennsylvanian Peninsula and eastern years ago Upper Peninsula, Michigan Basin. Peat deposits. Also oil and gas deposits 14. First Reptiles 300 million Pennsylvanian years ago 15. Gypsum and Limestone 320 million Mississippian deposited in shallow seas in years ago Lower Peninsula and eastern Upper Peninsula – Michigan Basin 16. First Sharks 350 million Devonian years ago Student No. Event Age Geologic Name Time Division 17. Hexagonaria coral deposits 360 million years Devonian (becomes MI state fossil aka ago Petosky Stone) in Michigan Basin 18. First Amphibians 370 million years Devonian ago 19. First Insects 400 million years Devonian ago 20. First Land Plants 420 million years Ordovician ago 21. -

University of Michigan University Library
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL.23, NO.5, p. 81-91, (4 pls.) JUNE 19, 1970 CORALS OF THE TRAVERSE GROUP OF MICHIGAN PART 13, HEXAGONARIA ERWIN C. STUMM MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Director: ROBERTV. KESLING The series of contributions from the Museum of Paleontology is a medium for the publication of papers based chiefly upon the collection in the Museum. When the number of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate papers may also be obtained. Correspondence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan 48104. VOLS.2-22. Parts of volumes may be obtained if available. Price lists available upon inquiry. 1. The rodents from the Hagerman local fauna, Upper Pliocene of Idaho, by Richard J. Zakrzewski. Pages 1-36, with 13 text-figures. 2. A new brittle-star from the Middle Devonian Arkona Shale of Ontario, by Robert V. Kesling. Pages 37-51, with 6 plates and 2 text-figures. 3. Phyllocarid crustaceans from the Middle Devonian Silica Shale of northwestern Ohio and southeastern Michigan, by Erwin C. Stumm and Ruth B. Chilman. Pages 53-71, with 7 plates and 4 text-figures. 4. Drepanaster wrighti, a new species of brittle-star from the Middle Devonian Arkona Shale of Ontario, by Robert V. Kesling. Pages 73-79, with 2 plates. -

CNIDARIA Corals, Medusae, Hydroids, Myxozoans
FOUR Phylum CNIDARIA corals, medusae, hydroids, myxozoans STEPHEN D. CAIRNS, LISA-ANN GERSHWIN, FRED J. BROOK, PHILIP PUGH, ELLIOT W. Dawson, OscaR OcaÑA V., WILLEM VERvooRT, GARY WILLIAMS, JEANETTE E. Watson, DENNIS M. OPREsko, PETER SCHUCHERT, P. MICHAEL HINE, DENNIS P. GORDON, HAMISH J. CAMPBELL, ANTHONY J. WRIGHT, JUAN A. SÁNCHEZ, DAPHNE G. FAUTIN his ancient phylum of mostly marine organisms is best known for its contribution to geomorphological features, forming thousands of square Tkilometres of coral reefs in warm tropical waters. Their fossil remains contribute to some limestones. Cnidarians are also significant components of the plankton, where large medusae – popularly called jellyfish – and colonial forms like Portuguese man-of-war and stringy siphonophores prey on other organisms including small fish. Some of these species are justly feared by humans for their stings, which in some cases can be fatal. Certainly, most New Zealanders will have encountered cnidarians when rambling along beaches and fossicking in rock pools where sea anemones and diminutive bushy hydroids abound. In New Zealand’s fiords and in deeper water on seamounts, black corals and branching gorgonians can form veritable trees five metres high or more. In contrast, inland inhabitants of continental landmasses who have never, or rarely, seen an ocean or visited a seashore can hardly be impressed with the Cnidaria as a phylum – freshwater cnidarians are relatively few, restricted to tiny hydras, the branching hydroid Cordylophora, and rare medusae. Worldwide, there are about 10,000 described species, with perhaps half as many again undescribed. All cnidarians have nettle cells known as nematocysts (or cnidae – from the Greek, knide, a nettle), extraordinarily complex structures that are effectively invaginated coiled tubes within a cell. -

Geology of Michigan and the Great Lakes
35133_Geo_Michigan_Cover.qxd 11/13/07 10:26 AM Page 1 “The Geology of Michigan and the Great Lakes” is written to augment any introductory earth science, environmental geology, geologic, or geographic course offering, and is designed to introduce students in Michigan and the Great Lakes to important regional geologic concepts and events. Although Michigan’s geologic past spans the Precambrian through the Holocene, much of the rock record, Pennsylvanian through Pliocene, is miss- ing. Glacial events during the Pleistocene removed these rocks. However, these same glacial events left behind a rich legacy of surficial deposits, various landscape features, lakes, and rivers. Michigan is one of the most scenic states in the nation, providing numerous recre- ational opportunities to inhabitants and visitors alike. Geology of the region has also played an important, and often controlling, role in the pattern of settlement and ongoing economic development of the state. Vital resources such as iron ore, copper, gypsum, salt, oil, and gas have greatly contributed to Michigan’s growth and industrial might. Ample supplies of high-quality water support a vibrant population and strong industrial base throughout the Great Lakes region. These water supplies are now becoming increasingly important in light of modern economic growth and population demands. This text introduces the student to the geology of Michigan and the Great Lakes region. It begins with the Precambrian basement terrains as they relate to plate tectonic events. It describes Paleozoic clastic and carbonate rocks, restricted basin salts, and Niagaran pinnacle reefs. Quaternary glacial events and the development of today’s modern landscapes are also discussed. -

Michigan Fossils Side by Side
MMiicchhiiggaann FFoossssiillss A2 - BRACHIOPOD, B2 - CRINOID pieces, A1 - BRACHIOPOD, B1 - CORAL or CHAIN invertebrate - invertebrate - invertebrate - CORAL, invertebrate - Mucrospifier profundus - Megistocrinus sp. ? - Pentamerus sp. - Cordell Halysites sp. - Cordell Silica formation - Alpena limestone - dolomite - Silurian age - dolomite - Silurian age - Devonian age - Devonian age - Alpena Chippewa Co. - 65 mm, Chippewa Co. - 110 mm, Washtenaw Co. - 50 mm, Co. - 30 mm (lower right) internal cast or steinkern siliceous replacement - calcite replacement - R. , calcite replacement - - R. Elowski R. Milstein Milstein GSD B4 - CORAL or A4 - CORAL or B3 - SNAIL or COLONIAL CORAL A3 - TRILOBITE, COLONIAL CORAL, GASTROPOD, Petoskey Stone, invertebrate - Phacops invertebrate - invertebrate - genus not invertebrate - rana - Silica formation - Syringopora sp. - Cordell determined - Cordell Hexagonaria percarinata Devonian age - dolomite - Silurian age - dolomite - Silurian age - - Alpena Limestone - Washtenaw Co. - 70 mm, s Chippewa Co. - 125 mm, s Chippewa Co. - 75 mm, Devonian age - calcite replacement - S. siliceous replacement - l internal cast or steinkern Charlevoix Co. - 200 mm, Wilson l R. Milstein - GSD calcite replacement - R. i i Milstein s s D1 - BANDED IRON C1 - MASTODON Tooth, C2 - EUCARYOTIC algae FORMATION (BIF), from D2 - PLANT root section, s vertebrate - Mammut filaments, plant - s fossils - result of plant - Stigmaria, genus americanum - Glacial Grypania spiralis - Grypania and others - not determined - Saginaw deposit - Quaternary age Negaunee Iron Formation Banded Iron Formation - Formation - o - 200 mm long, the - Precambrian age - o Precambrian age - Pennsylvanian age - “Michigan State Fossil” - Marquette Co. - large Marquette Co. - 600 mm, Eaton Co. - 150 mm in Central Michigan loop about 20 mm , F from the rock walkway at diameter, internal cast - F University Rowe Museum oldest macrofossil - GSD the - Eddy Discovery T. -

March 2020 Meetings and Field Trips Cancelled Until Further Notice Due to Covid-19 Concerns
Daisy Mountain Rockchips The purpose of Daisy Mountain Rock & Mineral Club is to promote and further an interest in geology, mineralogy, and lapidary arts, through education, field experiences, public service, and friendship. VOLUME 5, ISSUE 3 MARCH 2020 MEETINGS AND FIELD TRIPS CANCELLED UNTIL FURTHER NOTICE DUE TO COVID-19 CONCERNS A garden of corals in the now-buried (by lava) Kapoho Tidepools, Pahoa, Hawaii. This photo illustrates the colors missing from the fossil record. And it displays the potential growth forms -- many of which are related to the amount of turbulence in the water. Given species may be robust and massive in turbulent environments, and more delicately branching in quiet. Photo by Susan Celestian Daisy Mountain Rockchips March 2020 2 FOSSILS: PART V Kingdom: Animalia GRAPHITE Phylum: Cnidaria By Susan Celestian By Susan Celestian Graphite was known as plumbago, since 1739, but More complex than Porifera (sponges), the phylum was named for the Greek graphein (“to write”) by Cnidaria includes corals, sea anemones, sea Abraham Gottlieb Werner in 1789. It is one of pens, jellyfish, box jellies, hydrozoa (which include four minerals composed of carbon, the others the Portuguese Man O’War), stalked jellyfish, and being: diamond, chaoite, and lonsdaleite. other lesser known groups and species. Of these, Diamond -- the hardest natural substance known by far the majority in the fossil record are the on Earth -- has the same composition (but different corals, because they include species that create a crystal structure) as Graphite -- one of the softest hard calcium carbonate structure; while the others natural substances. are largely soft-bodied with no hard parts. -

Paleozoic Corals of Alaska
Paleozoic Corals of Alaska Geologic and Paleogeographic Setting of Paleozoic Corals in Alaska Ordovician, Silurian, and Devonian Corals of Alaska Carboniferous Corals of Alaska A Preliminary Report Stratigraphic Distribution of Permian Corals in Alaska GEOLOGICAL SURVEY PROFESSIONAL PAPER 823-A, B, C, D Paleozoic Corals of Alaska Geologic and Paleogeographic Setting of Paleozoic Corals in Alaska By MICHAEL CHURKIN, JR. Ordovician, Silurian, and Devonian Corals of Alaska By WILLIAM A. OLIVER, JR., CHARLES W. MERRIAM, and MICHAEL CHURKIN, JR. Carboniferous Corals of Alaska A Preliminary Report By AUGUSTUS K. ARMSTRONG Stratigraphic Distribution of Permian Corals in Alaska By CHARLES L. ROWETT GEOLOGICAL SURVEY PROFESSIONAL PAPER 823-A, B, C, D UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1975 UNITED STATES DEPARTMENT OF THE INTERIOR STANLEY K. HATHAWAY, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress Cataloging in Publication Data Main entry under title: Paleozoic corals of Alaska. (Geological Survey professional paper ; 823) CONTENTS: Churkin, M. Jr. Geologic and paleogeographic setting of Paleozoic corals in Alaska. Oliver, W. A. Jr., Merriam, C. W. and Churkin, M., Jr. Ordovician, Silurian, and Devonian corals of Alaska, [etc.] Includes bibliographies and indexes. Supt. of Docs, no.: I 19.16:823-A,B,C,D 1. Corals, Fossil. 2. Paleontology Paleozoic. 3. Paleiontology Alaska. I. Churkin, Michael, 1932- II. Series: United States. Geological Survey. Professional paper ; 823. QE778.P34 563'.6'09798 75-619102 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Stock Number 024-001-02666-0 CONTENTS [The letters in parentheses preceding the titles are those used to designate the chapters] Page (A) Geologic and paleogeographic setting of Paleozoic corals in Alaska, by Michael Churkin, Jr. -

The Petoskey Stone Some History, Lore and Facts About the "Petoskey Stone"
The Petoskey Stone Some history, lore and facts about the "Petoskey Stone" Wouldn't it be great if rocks could talk? For example, Today, Petoskey is a growing city with all of the through legend and facts, a Petoskey Stone could tell comforts of modern life and an appreciation of the past. you much about Michigan's history and geology. This is where Petoskey Stones are found. For those who look, Petoskey Stones are along the beaches, inland in Legend and history are often intertwined. Such is the gravel deposits, and sold in gift shops. case with the Petoskey Stone. The name Petoskey Stone likely came about because it was found and sold as The most often asked question is, "What is a Petoskey a souvenir from the Petoskey area. The name Petoskey Stone?" A Petoskey is a fossil colonial coral. These appears to have originated late in the 18th century. Its corals lived in warm shallow seas that covered Michigan roots stem from an Ottawa Indian legend. According to during Devonian time, some 350 million years ago. legend, a descendant of French nobility named Antoine Carre visited what is now the Petoskey area and became Almost a century after the founding of Petoskey, on a fur trader with the John Jacob Astor Fur Company. June 28, 1965, Governor George Romney signed a bill In time, he met and married an Ottawa Indian princess. that made the Petoskey Stone Michigan's official State Carre became known to the Indians as Neatooshing. Stone. It was fitting that Miss Ella Jane Petoskey, the Eventually he was adopted by the tribe and was made only living grand-child of Chief Petosegay, was present chief. -

Guide to Fossil Collecting in Minnesota
Educational Series - 1 Guide To Fossil Collecting in Minnesota Sarah Tufford and Rudolph Hogberg }lIN Minnesota Geological Survey qs 13 : J UNIVERSITY OF MINNESOTA First edition 1965 Minnesota Geological Survey Paul K. Sims, Director Minneapolis, Minnesota· 55455 Guide To Fosstl Collecting in Minnesota Sarah Tufford and Rudolph Hogberg Illustrations by Ann Cross Geologic Time Chart_Minnesota Time Era Period Events in Minnesota Characteristic Life 0 Continentol glociation Quaternary 20 ENO- followed b worm period Age ofMammals ZOIC 40 Tertiary No record in Minnesota 60 M Sea enters Minnesota from E Cretaceous 100 S West. Deposition of sediments. 0 Z Age of Reptiles 0 Jurassic I No record in Minnesota Triassic ~ C 200 Permian Age of Pennsylvanian No record in Minnesota C> Amphibians t), C> Mississippian ~ ~ ~ P Sea enters Minnesota from 300 Devonian ..... South. Deposition of sediments. AgeOfFIS~ C> A :@ l ~ Silurian No record in Minnesota Age of Corals ~ E 0 Age of Straight 40a Ordovician Seas cover Minnesota at intervals. Z ~PM~ 0 C Cambrian Deposition of sediments. Age of Trilab/~es 500 6 0 Lava flows and deposition of sediments. PRECAMBRIAN Deposition of iron-rich sediments. First record of life 4 Vz billion years lang Formation of mountains and igneous intrusions. Guide To Fossil Collecting in Minnesota Sarah Tufford and Rudolph Hogberg FOSSILS te 11 us what life was like on earth in ancient geologic time. A fossil clam, for example, lived on a sea bottom much as its modern relatives do. By finding many fossil clams, we can deter mine the extent of a prehistoric sea. Fossils also indicate the climates of the geologic past. -

Fossils 2019 Coaches Clinic Presentation (2020 Rules)
NCSO Institute Coaches Clinic 2019 FOSSILS Information for Coaches and Contestants Overview: Main ideas • Fossil identification and • Dating and the Geologic Time classification Scale • group morphology, adaptations • index fossils and correlation vs. • Prehistoric life radiometric dating • ecologic relationships, behaviors → • Important discoveries and trace fossils evolutionary events/transitions • habitat, environmental adaptations • e.g. discovery of transitional species • Geologic processes such as Tiktaalik, Archaeopteryx • • fossil formation (conditions, major events such as Cambrian processes), lagerstätten explosion, evolution of tetrapods, Permian extinction • sedimentary rocks, mineral/organic tissue composition Overview: Format • Emphasis on task-oriented activities • Rotating through stations (e.g. 20 stations, 2 minutes each) • Binder (≤ 2 inches this year) and Official Fossil List • Can’t usually remove materials from binder during competition • Magnifying glass • No guidebook The Fossil List • List with ~100 groups (will only need to identify groups on the list, but other groups may be used to illustrate key concepts) • Team members can divide up the fossil groups for depth • Changes this year • a few new genera (mostly dinosaurs, hominids) Order Tabulata (tabulate corals) • some groups marked for states/nationals only Genus Favosites • emphasis on intermediate levels of classification Genus Halysites* Order Rugosa (rugose corals) • e.g. corals and dinosaurs Genus Heliophyllum (horn coral) • form taxa for plants Genus -

GB28: Carbonate Platform Facies and Faunas of the Middle and Upper Devonian Cedar Valley Group and Lime Creek Formation, Norther
CARBONATE PLATFORM FACIES AND FAUNAS OF THE MIDDLE AND UPPER DEVONIAN CEDAR VALLEY GROUP AND LIME CREEK FORMATION, NORTHERN IOWA Iowa Geological and Water Survey Guidebook Series No. 28 Iowa Department of Natural Resources Richard Leopold, Director October 2008 COVER Lithographic plate of Idlewild Member of Lithograph City Formation at Lewis Quarry, immediately south of Osage, Mitchell County, Iowa. Beds marked “X” are those determined by A. Hoen & Co. of Baltimore to be suitable for use in lithography. Reproduced from Plate VIII in Iowa Geological Survey Annual Report for 1902. Printed in-house on recycled paper. CARBONATE PLATFORM FACIES AND FAUNAS OF THE MIDDLE AND UPPER DEVONIAN CEDAR VALLEY GROUP AND LIME CREEK FORMATION, NORTHERN IOWA Iowa Geological and Water Survey Guidebook Series No. 28 Prepared for the 69th Annual Tri-State and Great Lakes Section – SEPM Fall Field Conference Co-hosted by the Department of Earth Science, University of Northern Iowa and the Iowa Geological and Water Survey Prepared and led by John R. Groves1, James C. Walters1 and Jed Day2 with contributions by Rodney Hubsher1, Carl W. Stock3, Brian J. Witzke4 and Bill J. Bunker4 1 Department of Earth Science, University of Northern Iowa, Cedar Falls, IA 50614-0335 2Department of Geography & Geology, Illinois State University, Normal, IL 61790-4100 3Department of Geological Sciences, University of Alabama, Tuscaloosa, AL 35487-0338 4Iowa Geological and Water Survey, 109 Trowbridge Hall, Iowa City, IA 52242-1319 October 2008 Iowa Department of Natural Resources Richard Leopold, Director TABLE OF CONTENTS Overview of the Lithograph City Formation . 1 John R. Groves Quaternary geology of the fi eld trip area .