Silurian Rugose Corals of the Central and Southwest Great Basin
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

G. D. Eberlein, Michael Churkin, Jr., Claire Carter, H. C. Berg, and A. T. Ovenshine
DEPARTMENT OF THE INTERIOR GEOLOGICAL SURVEY GEOLOGY OF THE CRAIG QUADRANGLE, ALASKA By G. D. Eberlein, Michael Churkin, Jr., Claire Carter, H. C. Berg, and A. T. Ovenshine Open-File Report 83-91 This report is preliminary and has not been reviewed for conformity with U.S. Geological Survey editorial standards and strati graphic nomenclature Menlo Park, California 1983 Geology of the Craig Quadrangle, Alaska By G. D. Eberlein, Michael Churkin, Jr., Claire Carter, H. C. Berg, and A. T. Ovenshine Introduction This report consists of the following: 1) Geologic map (1:250,000) (Fig. 1); includes Figs. 2-4, index maps 2) Description of map units 3) Map showing key fossil and geochronology localities (Fig. 5) 4) Table listing key fossil collections 5) Correlation diagram showing Silurian and Lower Devonian facies changes in the northwestern part of the quadrangle (Fig. 6) 6) Sequence of Paleozoic restored cross sections within the Alexander terrane showing a history of upward shoaling volcanic-arc activity (Fig. 7). The Craig quadrangle contains parts of three northwest-trending tectonostratigraphic terranes (Berg and others, 1972, 1978). From southwest to northeast they are the Alexander terrane, the Gravina-Nutzotin belt, and the Taku terrane. The Alexander terrane of Paleozoic sedimentary and volcanic rocks, and Paleozoic and Mesozoic plutonic rocks, underlies the Prince of Wales Island region southwest of Clarence Strait. Supracrustal rocks of the Alexander terrane range in age from Early Ordovician into the Pennsylvanian, are unmetamorphosed and richly fossiliferous, and aopear to stratigraphically overlie pre-Middle Ordovician metamorphic rocks of the Wales Group (Eberlein and Churkin, 1970). -

Contents List of Illustrations LETTER OF
STATE OF MICHIGAN Plate IV. A. Horizontal and oblique lamination, Sylvania MICHIGAN GEOLOGICAL AND BIOLOGICAL SURVEY Sandstone......................................................................27 Plate IV. B. Stratification and lamination, in sand dune, Dune Publication 2. Geological Series 1. Park, Ind.........................................................................28 THE MONROE FORMATION OF SOUTHERN Plate V. Sand grains, enlarged 14½ times ............................31 MICHIGAN AND ADJOINING REGIONS Plate VI. Desert sand grains, enlarged 14½ times ................31 by Plate VII. Sylvania and St. Peter sand grains, enlarged 14½ A. W. Grabau and W. H. Sherzer times. .............................................................................32 PUBLISHED AS PART OF THE ANNUAL REPORT OF THE BOARD OF GEOLOGICAL AND BIOLOGICAL SURVEY FOR Figures 1909. Figure 1. Map showing distribution of Sylvania Sandstone. 25 LANSING, MICHIGAN WYNKOOP HALLENBECK CRAWFORD CO., STATE Figure 2. Cross bedding in Sylvania sandstone ....................27 PRINTERS Figure 3. Cross bedding on east wall of Toll’s Pit quarry ......28 1910 Figure 4. Cross bedding shown on south wall of Toll’s Pit quarry.............................................................................28 Contents Figure 5. Cross bedding on south wall of Toll’s Pit quarry in Sylvania sandstone. .......................................................28 Letter of Transmittal. ......................................................... 1 Figure 6. Cross bedding shown on south wall -

United States
DEPARTMENT OF THE INTERIOR BULLETIN OF THE UNITED STATES ISTo. 146 WASHINGTON GOVERNMENT Pit IN TING OFFICE 189C UNITED STATES GEOLOGICAL SURVEY CHAKLES D. WALCOTT, DI11ECTOK BIBLIOGRAPHY AND INDEX NORTH AMEEICAN GEOLOGY, PALEONTOLOGY, PETEOLOGT, AND MINERALOGY THE YEA.R 1895 FEED BOUGHTON WEEKS WASHINGTON Cr O V E U N M K N T P K 1 N T I N G OFFICE 1890 CONTENTS. Page. Letter of trail smittal...... ....................... .......................... 7 Introduction.............'................................................... 9 List of publications examined............................................... 11 Classified key to tlio index .......................................... ........ 15 Bibliography ............................................................... 21 Index....................................................................... 89 LETTER OF TRANSMITTAL DEPARTMENT OF THE INTEEIOE, UNITED STATES GEOLOGICAL SURVEY, DIVISION OF GEOLOGY, Washington, D. 0., June 23, 1896. SIR: I have the honor to transmit herewith the manuscript of a Bibliography and Index of North American Geology, Paleontology, Petrology, and Mineralogy for the year 1895, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. 1 BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINER ALOGY FOR THE YEAR 1895. By FRED BOUGHTON WEEKS. INTRODUCTION. The present work comprises a record of publications on North Ameri can geology, paleontology, petrology, and mineralogy for the year 1895. It is planned on the same lines as the previous bulletins (Nos. 130 and 135), excepting that abstracts appearing in regular periodicals have been omitted in this volume. Bibliography. The bibliography consists of full titles of separate papers, classified by authors, an abbreviated reference to the publica tion in which the paper is printed, and a brief summary of the con tents, each paper being numbered for index reference. -

Lines in Paradoxides Forchhammeri - Evidence of Lateral Inhibition
Spatial organization of tubercles and terrace lines in Paradoxides forchhammeri - evidence of lateral inhibition 0YVIND HAMMER Hammer, 0. 2000. Spatial organization of tubercles and terrace lines in Paradoxides forchhammeri - evidence of lateral inhibition. -Acta Palaeontologica Polonica 45,3, 251-270. Spatial statistics on the positions of trilobite tubercles indicate the existence of a develop- mental spacing mechanism. Similar spacing between sensory bristles, due to lateral inhi- bition, is well known in insects, and the genetic basis for these patterns has been thor- oughly studied. Tubercles (granules) in the Middle Cambrian trilobite Paradoxides forchhammeri are spaced out, but otherwise randomly positioned. Assuming that similar genetic principles are in operation for the positioning of peripheral neuronal elements in all arthropods, it can even be speculated that genes with functions similar to Delta, Notch, achaete and scute were active in trilobite cuticular patterning. Also, in f? forchhammeri, terrace lines (ridges) seem to display transitions into granulation, indicating that these two types of structure share an underlying pattern formation mechanism. Key words : Trilobites, tubercles, terrace lines, pattern formation, evolution, Notch. 0yvind Hammer [[email protected]],Paleontological Museum, University of Oslo, Sars gt. 1, 0562 Oslo, Norway. Introduction Trilobite tubercles (Starrmer 1980; Wilmot 1991; Whittington 1997) are cuticular structures that may have formed in connection with sensory devices such as setae or other mechano- or chemoreceptors. As such, they may be compared with sensilla in the cuticles of other arthropods (Starrmer 1980). In particular, the molecular basis for the development of spaced bristles in Drosophila has been the subject of extensive re- search (reviewed in Simpson et al. -

Morphology, Faunas and Genesis of Ordovician Hardgrounds from Southern Ontario, Canada
Palaeogeogmphy, Paheoclimatology, Pudoeoecology, 46 (1984): 233-290 Elsevier Science Publishers B.V., Amsterdam -Printed in The Netherlands MORPHOLOGY, FAUNAS AND GENESIS OF ORDOVICIAN HARDGROUNDS FROM SOUTHERN ONTARIO, CANADA CARLTON E. BRETT and MICHAEL E. BROOKFIELD' Depnrtmenl of Geological Sciences, University of Rochester, Rochester, NY 14627 (U.S.A,] Department v f Land Resources Sciences, GueIph University, Guelph, Ont. N.l G 2 W1 (Canada) {keceived May 19, 1983;revised version accepted March 13, 1984) ABSTRACT Brett, C. E. and Brookfield, M. E., 1984. Morphology, faunas and genesis of Ordovician hardgrounds from southern Ontario, Canada. Palaeogeogr., PaIaeoclirnatol., PalaeoecoI., 46: 233-290. We have used associations of different rnicrofacies to define facies (or microfaties associations) which fom reasonably welldefined sequences, which we infer, from amlo- gies with recent and ancient carbonate environments, to have been deposited in a shelf environment characterized by small-scde topographic differentiation into shoal, slope and basinal environments. Shoal environments are characterized by typically cross-bedded, well-sorted biocktic sands, with intershoal areas consisting oP interbedded bioclastic sands and heavily biotur- bated f iner-grained carbonates. Slope and "basinal" environments are typically represented by "proximal" and "distal" cycles respectively. These we compare with deposits of carbonate ramp bypass channels, and with the more thoroughly studied deep-water elastic submarine fans. Many of the strong variations in environmental energy in these proximal and distal cycles can be attributed ta migration of channels on the fans and to the effect of funnelling of storm surges dawn the channels, Mthough hardground morphology and faunas are mostly related to local effects rmch as intensity of scouring, time of exposure, topographic differentiation of the surface and other factors, differing hardground types tend to be found in different environments. -

Trilobites from the Silurian of New South Wales
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Fletcher, Harold O., 1950. Trilobites from the Silurian of New South Wales. Records of the Australian Museum 22(3): 220–233, plates xv–xvi. [27 January 1950]. doi:10.3853/j.0067-1975.22.1950.603 ISSN 0067-1975 Published by the Australian Museum, Sydney nature culture discover Australian Museum science is freely accessible online at http://publications.australianmuseum.net.au 6 College Street, Sydney NSW 2010, Australia TRILOBITES FROM THE SILURIAN OF NEW SOUTH WALES. By H. O. FLETCHER. Curator of Palaeontology, The Australian Museum. (Plates xv-xvi.) In this paper three new species of trilobites are described from a Lower Silurian horizon at Borenore, near Orange, New South Wales, as Encrinurus borenorensis, Phacops macdonaldi and Dicranogmus bartonensis. The genus Encrinurus Emmrich, 1844, is discussed and it is considered that the genus Gryptonymus Eichwald, 1825, is an abandoned name and cannot be used outside certain limits. Reference is made to the recorded Australian species of the family Lichidae and their geological age. The fossil material was originally found and forwarded to the Australian Museum by Mr. George McDonald, of "Rosyth", Borenore, on whose property the new horizon of fossils is situated. The author visited the locality later and collected additional specimens of all the described species. My thanks are due to Mr. McDonald for his assistance and interest, which have made possible the preparation of this paper. I am also indebted to Mr. F. Booker and Mr. L. Hall, of the Geological Survey of New South Wales, for assistance in determining the geological succession of the area. -
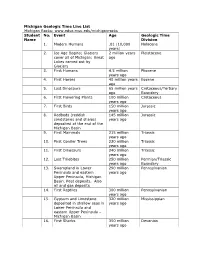
Michigan Geologic Time Line List Michigan Rocks: Student No
Michigan Geologic Time Line List Michigan Rocks: www.educ.msu.edu/michiganrocks Student No. Event Age Geologic Time Name Division 1. Modern Humans .01 (10,000 Holocene years) 2. Ice Age Begins; Glaciers 2 million years Pleistocene cover all of Michigan; Great ago Lakes carved out by Glaciers 3. First Humans 4.5 million Pliocene years ago 4. First Horses 40 million years Eocene ago 5. Last Dinosaurs 65 million years Cretaceous/Tertiary ago Boundary 6. First Flowering Plants 100 million Cretaceous years ago 7. First Birds 150 million Jurassic years ago 8. Redbeds (reddish 145 million Jurassic sandstones and shales) years ago deposited at the end of the Michigan Basin 9. First Mammals 215 million Triassic years ago 10. First Conifer Trees 230 million Triassic years ago 11. First Dinosaurs 240 million Triassic years ago 12. Last Trilobites 250 million Permian/Triassic years ago Boundary 13. Swampland in Lower 290 million Pennsylvanian Peninsula and eastern years ago Upper Peninsula, Michigan Basin. Peat deposits. Also oil and gas deposits 14. First Reptiles 300 million Pennsylvanian years ago 15. Gypsum and Limestone 320 million Mississippian deposited in shallow seas in years ago Lower Peninsula and eastern Upper Peninsula – Michigan Basin 16. First Sharks 350 million Devonian years ago Student No. Event Age Geologic Name Time Division 17. Hexagonaria coral deposits 360 million years Devonian (becomes MI state fossil aka ago Petosky Stone) in Michigan Basin 18. First Amphibians 370 million years Devonian ago 19. First Insects 400 million years Devonian ago 20. First Land Plants 420 million years Ordovician ago 21. -

Kyana Events -- Members Only! Rhabdosomes Including the Sicula and Spinose Thecae
http://www.amfed.org/sfms/index.html The KYANA Geological Society video produced by member Dave Shuffett can also be found on KET EncyloMedia internet site: http://www.ket.org/itvvideos/offering/science/ksroc.htm Electronic Edition of "LODESTAR" http://www.amfed.org/sfms/pdf/lodestar_may-june09.pdf MEETING TIME: 3rd Tuesday of each month - Social/Swap at 7:00 p.m. Meeting Starts at 7:30. Go To: http://www.kyanageo.org LOCATION: The Louisville Nature Center 3745 Illinois Avenue, Louisville, KY 40213 handicap accessible facility - visitors welcome http://www.louisvillenaturecenter.org DIRECTIONS: From the Watterson Expressway (I-264) -Take Exit 14 to Poplar Level Road North. Take Poplar Level Road to Trevilian Way (1.0 mi). Turn Right on to Trevilian Way. Take Trevilian Way to Illinois Avenue (0.2 mile) (to your left, Illinois Avenue is the first intersecting KYANA’s mission is to provide educational, road just before the Zoo entrance). recreational and social opportunities to those Turn left on to Illinois Avenue. interested in geological sciences and lapidary arts. Take Illinois Avenue to the Louisville Nature Center parking lot on your right (0.2 mile). Except for items specifically copyrighted by authors, other non-profit societies may use material in this newsletter. KYANA GEOLOGICAL SOCIETY Indiana and Kentucky / Volume 44, Number 9 THE GEMSCOOP c/o Sherry Lindle 6004 Highplace Dr Louisville, KY 40291 Revised July 7, 1999 at the AFMS Annual Meeting KYANA GEOLOGICAL SOCIETY Member of the The Southeast Federation of Mineralogical Societies, Inc. and American Federation of Mineralogical Societies “Code of Ethics” I will respect both private and public property and will do no collecting I will cause no willful damage to collecting materials and will take home only on privately owned land without the owner’s permission. -

University of Michigan University Library
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL.23, NO.5, p. 81-91, (4 pls.) JUNE 19, 1970 CORALS OF THE TRAVERSE GROUP OF MICHIGAN PART 13, HEXAGONARIA ERWIN C. STUMM MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Director: ROBERTV. KESLING The series of contributions from the Museum of Paleontology is a medium for the publication of papers based chiefly upon the collection in the Museum. When the number of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate papers may also be obtained. Correspondence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan 48104. VOLS.2-22. Parts of volumes may be obtained if available. Price lists available upon inquiry. 1. The rodents from the Hagerman local fauna, Upper Pliocene of Idaho, by Richard J. Zakrzewski. Pages 1-36, with 13 text-figures. 2. A new brittle-star from the Middle Devonian Arkona Shale of Ontario, by Robert V. Kesling. Pages 37-51, with 6 plates and 2 text-figures. 3. Phyllocarid crustaceans from the Middle Devonian Silica Shale of northwestern Ohio and southeastern Michigan, by Erwin C. Stumm and Ruth B. Chilman. Pages 53-71, with 7 plates and 4 text-figures. 4. Drepanaster wrighti, a new species of brittle-star from the Middle Devonian Arkona Shale of Ontario, by Robert V. Kesling. Pages 73-79, with 2 plates. -

CNIDARIA Corals, Medusae, Hydroids, Myxozoans
FOUR Phylum CNIDARIA corals, medusae, hydroids, myxozoans STEPHEN D. CAIRNS, LISA-ANN GERSHWIN, FRED J. BROOK, PHILIP PUGH, ELLIOT W. Dawson, OscaR OcaÑA V., WILLEM VERvooRT, GARY WILLIAMS, JEANETTE E. Watson, DENNIS M. OPREsko, PETER SCHUCHERT, P. MICHAEL HINE, DENNIS P. GORDON, HAMISH J. CAMPBELL, ANTHONY J. WRIGHT, JUAN A. SÁNCHEZ, DAPHNE G. FAUTIN his ancient phylum of mostly marine organisms is best known for its contribution to geomorphological features, forming thousands of square Tkilometres of coral reefs in warm tropical waters. Their fossil remains contribute to some limestones. Cnidarians are also significant components of the plankton, where large medusae – popularly called jellyfish – and colonial forms like Portuguese man-of-war and stringy siphonophores prey on other organisms including small fish. Some of these species are justly feared by humans for their stings, which in some cases can be fatal. Certainly, most New Zealanders will have encountered cnidarians when rambling along beaches and fossicking in rock pools where sea anemones and diminutive bushy hydroids abound. In New Zealand’s fiords and in deeper water on seamounts, black corals and branching gorgonians can form veritable trees five metres high or more. In contrast, inland inhabitants of continental landmasses who have never, or rarely, seen an ocean or visited a seashore can hardly be impressed with the Cnidaria as a phylum – freshwater cnidarians are relatively few, restricted to tiny hydras, the branching hydroid Cordylophora, and rare medusae. Worldwide, there are about 10,000 described species, with perhaps half as many again undescribed. All cnidarians have nettle cells known as nematocysts (or cnidae – from the Greek, knide, a nettle), extraordinarily complex structures that are effectively invaginated coiled tubes within a cell. -

XLVI. Notes on the Age of the Limestones of South Devonshire
[ 721 ] XLVI. Notes on the Age of the Limestones of South Devonshire. BY WILLIAM LONSDALE, P.G.S. [Read March 25, 1840.]* THE reasons which induced me to assume on zoological evidence, that the limestones of Southern Devon, between Dartmoor and the coast, would prove to be of the age of the old red sandstone, not having* been placed on record in a separate form, Mr. Murchison has begged me to lay before the Society a distinct notice of my claim to having been the first to propose the classifi cation recently put forth by Professor Sedgwick and himself; and in conse quence of an extension of those views to Belgium and the Boulonnais, Dr. Pitton has urged me to comply with the request. In the memoirsf in which Professor Sedgwick and Mr. Murchison announced the change in their classi fication of the older sedimentary strata of Devonshire and Cornwall, the clearest and amplest acknowledgements are made of my share in promoting the alteration; but as it is possible that some mistake may hereafter arise, re lative to the nature and limits of my suggestion, I conceive it is a duty to others as well as to myself to comply with the above request. I regret, how ever, that I cannot make this notice as satisfactory as I could wish, from the whole of the data not being accessible ; and I regret that my official duties have prevented me from bestowing upon it that time and thought and re search which ought to be evident in the shortest communications read to this Society. -

Geology of Michigan and the Great Lakes
35133_Geo_Michigan_Cover.qxd 11/13/07 10:26 AM Page 1 “The Geology of Michigan and the Great Lakes” is written to augment any introductory earth science, environmental geology, geologic, or geographic course offering, and is designed to introduce students in Michigan and the Great Lakes to important regional geologic concepts and events. Although Michigan’s geologic past spans the Precambrian through the Holocene, much of the rock record, Pennsylvanian through Pliocene, is miss- ing. Glacial events during the Pleistocene removed these rocks. However, these same glacial events left behind a rich legacy of surficial deposits, various landscape features, lakes, and rivers. Michigan is one of the most scenic states in the nation, providing numerous recre- ational opportunities to inhabitants and visitors alike. Geology of the region has also played an important, and often controlling, role in the pattern of settlement and ongoing economic development of the state. Vital resources such as iron ore, copper, gypsum, salt, oil, and gas have greatly contributed to Michigan’s growth and industrial might. Ample supplies of high-quality water support a vibrant population and strong industrial base throughout the Great Lakes region. These water supplies are now becoming increasingly important in light of modern economic growth and population demands. This text introduces the student to the geology of Michigan and the Great Lakes region. It begins with the Precambrian basement terrains as they relate to plate tectonic events. It describes Paleozoic clastic and carbonate rocks, restricted basin salts, and Niagaran pinnacle reefs. Quaternary glacial events and the development of today’s modern landscapes are also discussed.