Late Miocene) Transgression in the Northern Vienna Basin
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

SV EATON Ziersdorf Und FK Austria Wien
SVZ-News Aktuelles vom SV EATON Ziersdorf Ausgabe 30 März 2013 www.sv-ziersdorf.at Redaktionsschluss xx. März 2013 24. März: Eröffnung der Sportanlage Es sind nur noch wenige Stunden bis zur Eröffnung der Sportanlage – die Vorbereitungsarbeiten laufen auf Hochtouren! Dieser Event am Palmsonntag, 24. März 2013 soll einer der Höhepunkte in der 90-jährigen Ver- einsgeschichte des SV EATON Ziersdorf werden. Es haben sich eine Reihe von Ehrengästen angesagt, allen voran unser Ehrenpräsident, Landeshauptmann Dr. Erwin Pröll. Er wird die offizielle Eröffnung der Sportanlage vornehmen. Die Segnung der Anlage wird von Pfarrer Mag. Andreas Kalita vorgenommen. Weitere Programm- punkte beim Festakt sind neben der Begrüßung durch Obmann Manfred Binder die Reden von Bgm. ASVÖ- NÖ-Präsident Johann Gartner sowie NÖFV-Präsident Ludwig Binder. Für die musikalische Umrahmung des Festaktes wird die Trachtenkapelle Ziersdorf und Umgebung sorgen. Das anschließende Eröffnungsspiel zwischen SV EATON Ziersdorf und FK Austria Wien, dem überlegenen Tabellenführer der Bundesliga, soll sowohl für die Zuschauer als auch für die Spieler des SV EATON Ziersdorf zu einem unvergesslichen Erlebnis werden. Bei der Eröffnungsfeier wird der SV EATON Ziersdorf eine Charity-Aktion zugunsten der Katastrophenhilfe österr. Frauen durchführen. Präsidentin dieses Vereines ist Elisabeth Pröll, die Gattin des Landeshauptmannes. Nach dem Eröffnungsspiel stehen die Spieler von FK Austria Wien für eine Autogrammstunde zur Verfügung. Stehplatzkarten sind an der Tageskasse zum Preis von € 12,-- (für Erwachsene) bzw. € 6,-- (für Jugendliche zw. 10 und 15 Jahren) erhältlich. Kinder bis 10 Jahren haben freien Eintritt. In den letzten Wochen wurde vor allem auf die Gestaltung der Innenräume – insbeson- dere der Kantine, des VIP-Raumes und der Gangbereiche – das Augenmerk gelegt. -

Gemeindenachrichten Dezember 2018
der Marktgemeinde Nappersdorf-Kammersdorf Ausgabe 3/2018 Ausgabe 1/2016 Dürnleis Haslach Kammersdorf Kleinsierndorf Kleinweikersdorf Nappersdorf Haslach Kammers dorf Kleinsiern dorf Kleinweik ersdorf Nappersd orf Sehr geehrte Gemeindebürgerinnen und Gemeindebürger! Liebe Jugend! Ein klimatisch sehr warmes Jahr mit wenigen Niederschlägen neigt sich zu Ende. Zum Glück blieb unsere Gemeinde von Umweltkatastrophen verschont. Ich möchte mich herzlich bei den Gemeinderäten, den Ortsvorstehern, dem Gemeindeteam und allen Helfern - Vereinen oder Privatpersonen, die in Mit den besten Weihnachtsgrüßen irgendeiner Weise die Gemeinde mit ihrer Tatkraft Ihr Bürgermeister Gottfried Pompe unterstützt haben, oder durch ihren Einsatz lebenswerter gemacht haben - bedanken. Ohne deren freiwillige Leistungen wäre vieles nicht möglich gewesen. Impressum: Hersteller, Herausgeber, Redaktion: Marktgemeinde Nappersdorf-Kammersdorf, 2033 Kammersdorf, Kammersdorf 58 Im Namen des Gemeinderates und der Telefon: +43 2953 2314, Fax+43 2953 2314-15 Gemeindebediensteten wünsche ich Ihnen ein Homepage: www.nappersdorf-kammersdorf.gv.at UID: ATU 16228909, DVR.: 0002798 frohes Weihnachtsfest im Kreise Ihrer Familie und Verlags- und Herstellungsort: 2033 Kammersdorf 58 viel Glück und Gesundheit für das neue Jahr! Für den Inhalt verantwortlich: Bürgermeister Gottfried Pompe, 2033 Kammersdorf 58 GemeindePROJEKTE Betonspurwege In einem Pilotprojekt wurden in Nappersdorf und Kleinsierndorf Betonspurwege errichtet. Unter der Mithilfe der Landwirte und einem speziellen Betonfertiger -

Industrie + Gewerbe Landwirtschaft Und Tourismus Interkommunale Kooperationen Das Tor Zur Wirtschaft
– Standort Ziersdorf – INDUSTRIE + GEWERBE DAS TOR ZUR WIRTSCHAFT Optimaler Standort für Gewerbe- und Der Wirtschaftspark Schmidatal-Manhartsberg Industriebetriebe. Profitieren Sie von liegt im Herzen des Weinviertels – hier treffen sofort verfügbaren gewidmeten und aufgeschlossene Gewerbegründe mit Breitband- vollaufgeschlossenen Gewerbegründen. anschluss auf Kleinkunst und Kellergassen. Die Anbindung an leistungsfähige Straßen > HOHE LEBENSQUALITÄT und dem Schienenverkehr bietet beste > AUSGEZEICHNETE STANDORTQUALITÄT Infrastruktur und perfekte Logistik. > AUSGEZEICHNETES ARBEITSKRÄFTEPOTENZIAL > INTERESSANTER ABSATZMARKT LANDWIRTSCHAFT IM UMFELD UND TOURISMUS > 50 KM ZUR MILLIONENSTADT WIEN Herausragende Qualität der Region mit Authentizität in der Landwirtschaft, Gesamtangebote in der Erlebnis- Wien – B4-Umfahrung Ziersdorf – Horn gastronomie, von Produktion und Ver- BB ! edelung bis zum Konsum. Erreichen Sie Ra Knoten d l b Ziersdorf r neue Zielgruppen im Bereich Tourismus. u e n Mitt n – K r em s e BB rs t ra BB s s e – Z i e ERWEITERUNG MÖGLICH r sd INTERKOMMUNALE o BB r f BS KOOPERATIONEN BS BB Bauland, Betriebsgebiet VP BS Bauland, Sondergebiet Gastronomie VP Private VF Tankstelle Die Betreibergemeinden des Wirt- Gfrei Grünland-Freihaltefläche frei Gfrei BB verkauft schaftsparks setzen im Interesse der regionalen Lebensqualität Kooperations- > Expansionsfläche: 17-19 Hektar möglich projekte um. Mit dem regionalen Post- > Kaufpreis: € 12,80 bis € 14,80 pro m2 verteilerzentrum und dem Abfallzentrum > Techn. Infrastruktur: Strom, Kanal,Znaim Trinkwasser, Gmünd wurden sichtbare Zeichen gesetzt. Breitbandinternet, Telekommunikation,Retz Kläranlage Pulkau Horn > UnternehmenEggenburg vor Ort: SBI Produktion techn. Laa an der Thaya Anlagen GmbH. & Co KG, Post Zustellbasis, Abfallverband Sitzendorf Zwettl Maissau Limberg Ravelsbach Hollabrunn ZIERSDORF Ernstbrunn Glaubendorf Hohenwarth Heldenberg Goßweikersdorf Langenlois Schmid Krems Kirchberg 4 a Stockerau Donau A22 3 Tulln WIEN anz-Josefs-Bahn St. -

DIE BÖDEN DES WESTLICHEN WEINVIERTELS 97-105 ©Geol
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Arbeitstagung der Geologischen Bundesanstalt Jahr/Year: 1999 Band/Volume: 1999 Autor(en)/Author(s): Stich Robert Artikel/Article: DIE BÖDEN DES WESTLICHEN WEINVIERTELS 97-105 ©Geol. Bundesanstalt, Wien; download unter www.geologie.ac.at Arbeitstagung Geologische Bundesanstalt 1999 • Retz - Hollabrunn Beiträge DIE BÖDEN DES WESTLICHEN WEINVIERTELS Robert STICH Das flachwellige Hügelland des Weinviertels im Nordosten Niederösterreichs wird durch die Waschbergzone in das östliche Weinviertel mit dem Inneralpinen Wiener Becken und das westliche Weinviertel geteilt. Letzteres gehört zur Molassezone und wird von tertiären Tonen, Sanden, Mergeln und Schottern (z.B. Hollabrunner Schotter aus dem Pannonium), z.T. unter Lößbedeckung aufgebaut. Das westliche Weinviertel ist im Hauptproduktionsgebiet „Nordöstliches Flach- und Hügelland" in einem Seehöhenbereich von 195 m bis 450 m gelegen und umfaßt als Kleinproduktionsge biet die Landschaftsräume des Wagram, des Manhartsberges, des Schmidatales, des Pul kautales, des Pulkau-Retzer Weinlandes und der Hohenwarther Platte (WAGNER, 1990). Auf grund der Thematik der Arbeitstagung sollen aber hier die Böden des Retzer und Holla brunner Raumes dargestellt werden. Hier dominieren die Weinbaubetriebe, es sind jedoch auch viele "gemischte Landwirtschaftsbe triebe" und Marktfruchtbetriebe vorhanden, vor allem in den Grenzbereichen zum Hollabrunn- Mistelbacher -

Documentation Austrian Wine 2006
DOCUMENTATION AUSTRIAN W INE 2006 Table of contents 1 Austria œ the wine country 1.1 Austria‘s wine-gr wing regi ns and wine-gr wing areas 1 1.2 Grape varieties in Austria 5 1.2.1 Breakd wn by share of area in percent 5 1.2.2 Grape varieties - Brief descripti n .1 1.2.. Devel pment f the area under cultivati n until 1111 .5 1.. Devel pment of the climate 1161-2002 .6 1.2 W ine-gr wers in Austria - A current overall view .1 1.5 The 2006 harvest 22 1.6 The 2005 vintage 25 1.3 Brief characterisati n of the vintages 2002 back t 1160 23 1.8 Assessment of the 2005-111. vintages 55 2 The Austrian wine industry 2.1 Ec n mic imp rtance of the wine industry in Austria 56 2.2 The harvest 2006 (Status Oct ber 20066 51 2.. 7arvests 1160-2005 61 2.2 8ualit9tswein (8uality wine6 in Austria 2005 65 2.5 Austria‘s wine supply 2005 68 2.6 Devel pment f grape and wine prices 31 2.3 General regulati ns f r wine pr ducti n 32 2.8 EU-Measures f r the Restructuring and C nversi n of Vineyards 32 2.1 The Austrian W ine B ards 80 2.10 The :alue f Origin 8. 2.11 DAC: the l gical key t Austrian wine 82 2.12 8uesti ns and Answers - a Guide 86 3 The Austrian market ..1 C nsumpti n of D mestic Wine and Sparkling W ine 81 ..1.1 C nsumpti n of D mestic Wine 81 ..1.2 D mestic C nsumpti n f Sparkling Wine 1110-2005 10 ..1. -

Transgression. Foraminiferal and Ostracod Assemblages from an Incised Valley Fill in the Molasse Basin of Lower Austria
Austrian Journal of Earth Sciences Volume 102/2 Vienna 2009 The initial phase of the early Sarmatian (Middle Miocene) transgression. Foraminiferal and ostracod assemblages from an incised valley fill in the Molasse Basin of Lower Austria. Holger GEBHARDT*), Irene ZORN & Reinhard ROETZEL KEYWORDS paleoenvironmental interpretation Middle Miocene Lower Austria foraminifera Geologische Bundesanstalt, Neulinggasse 38, 1030 Wien, Austria; Sarmatian ostracods *) Corresponding author, [email protected] Badenian Abstract In order to document the foraminiferal and ostracod faunal changes between Badenian and Sarmatian strata, we investigated sam- ples from a sediment core drilled at Hollabrunn in Lower Austria. Lower Badenian strata show typical normal marine, outer shelf to upper bathyal foraminferal and ostracod assemblages also known from other sections in the Molasse and Vienna Basins. The co- occurrence of Praeorbulina glomerosa circularis and Orbulina suturalis indicates a lower Badenian age for these samples, corres- ponding to the mid Langhian Zone M6. The investigated core lacks middle to upper Badenian strata. We were able to identify the Sarmatian samples by 1) index species and 2) by apparently unique autochthonous microfossil assemblages. We found the Sarma- tian foraminiferal index species Nonion bogdanowiczi and Anomalinoides dividens in samples from the upper part of the core and did not find foraminifers or ostracods belonging to lower Sarmatian Elphidium reginum Zone or younger (eco-) Zones. Consequently, the drilled Sarmatian samples belong to the basal Sarmatian Anomalinoides dividens (eco-) Zone. The monospecific occurrence of Au- bignyna perlucida in one older sample, however, suggests an extreme, most likely brackish environment with very different condi- tions from those prevailing earlier during the Badenian. -

Programm 2021
Tal der Vielfalt. Programm 2021 1 Tal der Zukunft. In der aktuellen Zeit spürt, liest und redet man über notwendige und vor allem erwünschte Veränderungen im Denken und Handeln gegenüber unserem Naturerbe. Das vergangene Jahr hat gezeigt, dass der Bedarf und das Verständnis für den Erhalt der ursprünglichen Natur sehr vielen Menschen wichtig sind. Wir machen uns mehr Gedanken zu mehr Nach- haltigkeit im täglichen Leben, wir diskutieren über die Einsparung von CO2, es gibt junge, glo- bale Bewegungen wie die „Fridays for Future“ und es werden zukunftsweisende Abkommen wie der EU Green Deal geschlossen. Wir genießen es immer mehr, uns in der Natur zu be- wegen und bringen ihr mehr Wertschätzung entgegen – wir erleben den Beginn einer großen Veränderung. Darum stellen wir unser Jahresthema „Nature for Future“ an den Beginn einer Dekade des Wandels, der Anpassung und der Vielfalt. Schutzgebiete wie der Nationalpark Thayatal stellen dabei ganz besondere Kristallisations- punkte dar. Einerseits können hier die Prozesse weitestgehend ungestört ablaufen, die enor- me Vielfalt hat einen Ausgangspunkt und das weite Spektrum an Lebensräumen bietet Raum für Anpassungen an Veränderungen wie etwa den Klimawandel. Andererseits finden wir als Menschen einen wilden Erholungsraum, der unserem Wohlbefinden Vorschub leistet. Ganz im Sinne des Jahresthemas erscheint das vorliegende Programmheft in neuem Design und wir hoffen damit Lust auf die Teilnahme bei einer unserer vielfältigen Exkursionen und Veran- staltungen oder einen individuellen Ausflug in den „Green Canyon“ des Waldviertels zu ma- chen. Somit laden wir Sie mit großer Freude ein, im vorliegenden Programmheft zu schmö- kern und mit uns in die wilde Naturlandschaft des Thayatals einzutauchen! Christian Übl, BSc Mag. -
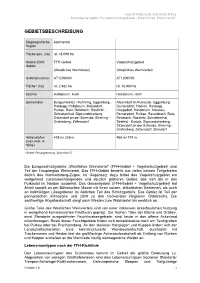
Gebietsbeschreibung
HAUPTREGION WEINVIERTEL Managementplan Europaschutzgebiete „Westliches Weinviertel“ GEBIETSBESCHREIBUNG Biogeografische kontinental Region Fläche ges. (ha) rd. 18.099 ha Natura 2000- FFH-Gebiet Vogelschutzgebiet Gebiet (Westliches Weinviertel) (Westliches Weinviertel) Gebietsnummer AT1209A00 AT1209V00 Fläche* (ha) rd. 2.982 ha rd. 16.904 ha Bezirke Hollabrunn, Horn Hollabrunn, Horn Gemeinden Burgschleinitz - Kühnring, Eggenburg, Alberndorf im Pulkautal, Eggenburg, Hardegg, Hollabrunn, Meiseldorf, Guntersdorf, Hadres, Hardegg, Pulkau, Retz, Retzbach, Röschitz, Haugsdorf, Hollabrunn, Maissau, Schrattenthal, Sigmundsherberg, Pernersdorf, Pulkau, Ravelsbach, Retz, Sitzendorf an der Schmida, Straning - Retzbach, Röschitz, Schrattenthal, Grafenberg, Zellerndorf Seefeld - Kadolz, Sigmundsherberg, Sitzendorf an der Schmida, Straning - Grafenberg, Zellerndorf, Ziersdorf Höhenstufen 438 m/ 226 m 468 m/ 191 m (max./min. m Höhe) * Quelle: Feinabgrenzung, Stand Mai 07 Die Europaschutzgebiete „Westliches Weinviertel” (FFH-Gebiet + Vogelschutzgebiet) sind Teil der Hauptregion Weinviertel. Das FFH-Gebiet besteht aus vielen kleinen Teilgebieten östlich des Manhartsberg-Zuges. Im Gegensatz dazu bildet das Vogelschutzgebiet ein weitgehend zusammenhängendes und deutlich größeres Gebiet, das sich bis in das Pulkautal im Norden ausdehnt. Das Gesamtgebiet (FFH-Gebiet + Vogelschutzgebiet) hat Anteil sowohl an der Böhmischen Masse mit ihren sauren, silikatischen Gesteinen, als auch an kalkhältigen Lössgebieten im östlichen Teil des Schutzgebiets. Das Gebiet ist -

Freiberuflichen Liste Hollabrunn
Physio Austria Niederösterreich http://www.physioaustria.at/niederoesterreich | [email protected] Stand per 19.07.2021 Irmtraud Lenius, MSc. Hollabrunn Badhausgasse 1 2020 Hollabrunn Alberndorf im Pulkautal Praxis rollstuhlgerecht T 02952 30505 Daniela Reinthaler-Scherz M 0699 1139 1895 Bahnstraße 193 F 02952 30505 2054 Alberndorf [email protected] M 0664 1627233 http://www.giz.co.at F 01 [email protected] Ilse Mühlparzer-Lebic Badhausgasse 1 Hollabrunn 2020 Hollabrunn Praxis rollstuhlgerecht Sabine Ingrid Gruber T 02952 30505 Wiener Straße 236 M 0664 1411765 2014 Breitenwaida [email protected] Praxis rollstuhlgerecht http://www.giz.co.at T 02954 3500 F 02954 3500 Maria Schmoll Badhausg. 1 Gerda Baar 2020 Hollabrunn Kapuzinerstraße 31 T 02952 30505 2020 Hollabrunn M 0676 6348614 T 02952 20718 F 02952 30505 M 0680 3058548 [email protected] [email protected] http://www.giz.co.at Renate Gallei Theresa Veigl, B.Sc. Badhausgasse 1 Pfarrgasse 4 2020 Hollabrunn 2020 Hollabrunn Praxis rollstuhlgerecht Praxis rollstuhlgerecht Hausbesuche Hausbesuche M 0681 81211897 T +436649125251 http://www.giz.co.at [email protected] http://www.theranova.at Christoph Gurtner Mitterhausergasse 17 Nappersdorf-Kammersdorf 2020 Hollabrunn Hausbesuche Daniela Gritsch T 0664/1617707 Nr. 222 M 0664/1617707 2033 Kammersdorf [email protected] Praxis rollstuhlgerecht Hausbesuche Patrick Kyncl, Bsc T +43 Badhausgasse 17 M 06769339693 2020 Hollabrunn [email protected] M 0680 4456638 https://www.physio-gritsch.at [email protected] http://www.physio-kyncl.at Physio Austria Niederösterreich http://www.physioaustria.at/niederoesterreich | [email protected] Stand per 19.07.2021 Pernersdorf Ziersdorf Johanna Schöfmann, B.Sc. -

Palaeontological Highlights of Austria
© Österreichische Geologische Gesellschaft/Austria; download unter www.geol-ges.at/ und www.biologiezentrum.at Mitt. Österr. Geol. Ges. ISSN 0251-7493 92 (1999) 195-233 Wien, Juli 2000 Palaeontological Highlights of Austria WERNER E. PILLER1, GUDRUN DAXNER-HÖCK2, DARYL P DOMNING3, HOLGER C. FORKE4, MATHIAS HARZHAUSER2, BERNHARD HUBMANN1, HEINZ A. KOLLMANN2, JOHANNA KOVAR-EDER2, LEOPOLD KRYSTYN5, DORIS NAGEL5, PETER PERVESLER5, GERNOT RABEDER5, REINHARD ROETZEL6, DIETHARD SANDERS7, HERBERT SUMMESBERGER2 28 Figures and 1 Table Introduction Besides Zeapora gracilis, distinguished by large rounded cortical filaments, Pseudolitanaia graecensis and Pseu The oldest known fossils in Austria date back into the dopalaeoporella lummatonensis occur (Fig. 3). Pseudolitan Ordovician. From this time on a broadly continuous fossil aia graecensis is built up of straight thalli containing club- record is preserved up to the Holocene. Since an encyclo shaped filaments and Pseudopalaeoporella lummatonensis paedic or monographic presentation is impossible within is characterized by a typically poorly-calcified medullar this volume, nine case studies of different stratigraphic lev zone and delicate cortical filaments. els (Fig. 1) were selected to call attention to this remarkably There are two localities known with autochthonous algal good fossil documentation. These case studies include occurrences in the Graz Palaeozoic. One is characterized records on invertebrate fossils from several time slices from by Pseudopalaeoporella lummatonensis with dispersed the Late Palaeozoic to the Miocene, as well as on verte thalli of Pseudolitanaia. Contrary to all expectations, these brates from the Miocene and Pleistocene and on plant algae are found in marly lithologies suggesting very bad fossils from the Devonian and Early Miocene. This selection environmental conditions for photoautotrophic organisms. -

Sektion Austria Alpenvereinshaus Telefon 01/513 10 03 A-1010 Wien, Rotenturmstraße 14 Telefax 01/513 10 03/17
Sektion Austria Alpenvereinshaus Telefon 01/513 10 03 A-1010 Wien, Rotenturmstraße 14 Telefax 01/513 10 03/17 http://www.alpenverein.at/austria e-mail: [email protected] Infoblatt Drosendorf und Umgebung Treffpunkt: Samstag, 4.8.2018 10:55 Uhr Bahnhof Drosendorf Anfahrt: Bahn oder Fahrgemeinschaften Rückkehr: Gegen 21:00 Tour: Von Drosendorf die Thaya am linken Ufer entlang flussabwärts, dann Aufstieg zur Ruine der gotischen Gaberkirche. Weiter durch das Tal des Gaberbachs nach Unterthürnau und am rechten Thayaufer über die Hubertuskapelle zurück nach Drosendorf. Der Stadtmauer entlang umrunden wir die Altstadt und bewundern das Häuser-Ensemble im Zentrum. Charakter: Einfache, aber ausgedehnte Wanderung durch Feld und Wald. Gehzeit: Circa 6 Stunden (inkl. Pause(n)). Ausrüstung: Der Witterung angepasste Wanderbekleidung und bequemes Schuhwerk, Regenschutz. Unterwegs keine Einkehr geplant, Jause und Getränk bitte mitnehmen! Karte: ÖK BMN Nr. 8 Führungsbeitrag: € 8,50 Bahn Retz-Drosendorf und retour mit NÖ-Card gratis! Information: 0664 415 24 34 Auf eine schöne Tour freuen sich Arnold und Elisabeth Geführte Touren der Sektion Austria – die Gründersektion des OEAV Sektion Austria Alpenvereinshaus Telefon 01/513 10 03 A-1010 Wien, Rotenturmstraße 14 Telefax 01/513 10 03/17 http://www.alpenverein.at/austria e-mail: [email protected] Hinfahrt: Station Datum Zeit Prognose Steig Verkehrsmittel Wien Liesing Bahnhof 04.08.2018 ab 07:36 3 R 2210 Wien Meidling Bahnhof ab 07:44 3 Wien Matzleinsdorfer Platz (ÖBB) ab 07:47 -

Historisches Ortslexikon
Historisches Ortslexikon Statistische Dokumentation zur Bevölkerungs- und Siedlungsgeschichte NIEDERÖSTERREICH 2. Teil Hollabrunn, Horn, Korneuburg, Krems (Land), Lilienfeld, Melk Datenbestand: 31.8.2016 1 BEZIRK HOLLABRUNN Gründungsbestand: über 3300 Urlehen (ohne abgekommene Siedlungen). - 1590: *6688, 1751: 8138, 1783/87: 9729-*59.900, 1794: 10.029-*58.500, 1795: *9873, 1822: 11.144, 1830: *11.640- 73.407, 1846: 11.884-76.857, 1851: 12.159-74.520, 1869: 13.082-71.791, 1880: 13.561-75.879, 1890: 13.939-77.584, 1900: 14.472-77.393, 1910: 15.126-76.688, 1923: 15.682-73.359, 1934: 16.660-72.202, 1939: 70.871, 1951: 16.802-67.637, 1961: 17.516-58.752, 1971: 18.731-54.963, 1981: 20.863-50.262, 1991: 22.297-49.066, 2001: 24.200-50.062, 2011: 24.893-50.301, 2012: 50.174, 2013: 50.065, 2014: 50.232, 2015: 50.617. Gründungsbestand: HR aus 120 Dorfsiedlungen (samt Stadt Hollabrunn, Märkte Göllersdorf und Sitzendorf an der Schmida, Repr. 78 % der H 1590): Gründungsanlagen etwa 2630 (Durchschnitt 22), 1590: 5247 H. Gemeinden: Q: Bezirk Hollabrunn (Bezemek Ernst - Rosner Willibald (Hg.), Vergangenheit und Gegenwart. Der Bezirk Hollabrunn und seine Gemeinden, 1993). – Koller Ludwig, Heimatbuch des Bezirkes Hollabrunn, 1. Teil, 1949. – 1497 (Teilungsurbare der Herrschaften Maissau und Sonnberg): Feigl Helmuth – Stockinger Thomas, Die Urbare der Herrschaften Maissau und Sonnberg anlässlich der Teilung des Erbes nach Georg von Eckartsau im Jahre 1497, Fontes Rerum Austriacarum, 3.Abt., Fontes Iuris 20, 2008. - 1499: Urbar Eggenburg, HKA Urbar 947. Ehem. Gerichtsbezirk Haugsdorf 1590: 640, 1751: 1056, 1783/86: 1316-9338, 1794: 1433-9340, 1795: 1449, 1822: 1807, 1830: 1903-13.370, 1846: 2006-13.903, 1851: 2035-14.271, 1857: 2115-13.769, 1869: 2313-13.473, 1880: 2482-14.454, 1890: 2645-15.509, 1900: 2751-14.967, 1910: 2894-14.395, 1923: 3002- 13.411, 1934: 3108-13.023, 1939: 12.588, 1951: 3095-11.659, 1961: 3106-9514, 1971: 3178- 8577, 1981: 3401-7498, 1991: 3544-6775, 2001: 3857-6810, 2011: 3952-6565, 2012: 6492, 2013: 6465, 2014: 6476, 2015: 6502.