Cannabinoids in the Pathophysiology of Skin Inflammation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Marinol Cannabidiol C21H30O2 Trade Name
Cannabinoids are a group of terpenophenolic compounds secreted by Cannabis flowers that provide relief from a wide array of symptoms including, pain, nausea, and inflammation. They operate by imitating the body’s natural endocannabinoids, which activate to maintain internal stability and overall health. When consumed, cannabinoids bind to receptor sites throughout the brain (CB1 receptors) and body (CB2 receptors). Different cannabinoids have different effects based on their binding affinity for each receptor. By targeting specific cannabinoids at these receptors, different types of relief can be achieved. Presently, there are at least 113 different cannabinoids isolated from Cannabis—each exhibiting varied effects. THC Tetrahydrocannabinol C21H30O2 Trade name: Marinol Legal Status: US – Schedule I, Schedule II (as Cesamet), Schedule III (as Marinol) OH CA – Schedule II UK – Class B AU – S8 (controlled) H Psychoactive Tetrahydrocannabinol (THC) is typically the most abundant cannabinoid present in cannabis products on the market today. THC has very high psychoactive characteristics and is associated with the ‘high’ and euphoria experienced when using cannabis products. When smoked or ingested, THC binds to cannabinoid receptors throughout the body and affects memory, O coordination, concentration, pleasure, and time perception. H Medicinal Benefits Analgesic • Anti-nauseant • Appetite Stimulant Reduces Glaucoma Symptoms Sleep Aid • Reduces Anxiety and PTSD Symptoms CBD Cannabidiol C21H30O2 Trade name: Epidiolex OH Legal Status: US – Schedule I CA – Schedule II UK – POM (Perscription only) H AU – S4 (Perscription only) non-psychoactive Cannabidiol (CBD) is a major phytocannabinoid and accounts for up to 40% of the plant’s extract. Due to its lack of psychoactivity and HO non-interference with motor and psychological functions, it is a leading candidate for a wide variety of medical applications. -

Recent Developments in Cannabis Chemistry
Recent Developments in Cannabis Chemistry BY ALEXANDER T. SHULGIN, Ph.D. The marijuana plant Cannabis sativa contains a bewildering Introduction array of organic chemicals. As is true with other botanic species, there are representatives of almost all chemical classes present, including mono- and sesquiterpenes, carbohy- drates, aromatics, and a variety of nitrogenous compounds. Interest in the study of this plant has centered primarily on the resinous fraction, as it is this material that is invested with the pharmacological activity that is peculiar to the plant. This resin is secreted by the female plant as a protective agent during seed ripening, although it can be found as a microscopic exudate through the aerial portions of plants of either sex. The pure resin, hashish or charas, is the most potent fraction of the plant, and has served as the source material for most of the chemical studies. The family of chemicals that has been isolated from this source has been referred to as the cannabinoid group. It is unique amongst psychotropic materials from plants in that there are no alkaloids present. The fraction is totally nitro- gen-free. Rather, the set of compounds can be considered as analogs of the parent compound cannabinol (I), a fusion product of terpene and a substituted resorcinol. Beyond the scope of this present review are such questions as the distribution of these compounds within the plant, the bo- tanic variability resulting from geographic distribution, the diversity of pharmacological action assignable to the several Reprinted from Journal of Psychedelic Drugs, vol. II, no. 1, 197 1. 397 398 Marijuana: Medical Papers distinct compounds present, and the various preparations and customs of administration. -

The Handbook of Cannabis Therapeutics: from Bench to Bedside
Handbook of Cannabis Therapeutics From Bench to Bedside 9780789030979 Handbook of Cannabis Therapeutics From Bench to Bedside Size: 212 x 152mm Spine size: 26 mm Color pages: Binding: Paperback THE HAWORTH PRESS® Haworth Series in Integrative Healing Ethan Russo Editor The Last Sorcerer: Echoes of the Rainforest by Ethan Russo Professionalism and Ethics in Complementary and Alternative Medicine by John Crellin and Fernando Ania Cannabis and Cannabinoids: Pharmacology, Toxicology, and Therapeutic Potential by Franjo Grotenhermen and Ethan Russo Modern Psychology and Ancient Wisdom: Psychological Healing Practices from the World’s Religious Traditions edited by Sharon G. Mijares Complementary and Alternative Medicine: Clinic Design by Robert A. Roush Herbal Voices: American Herbalism Through the Words of American Herbalists by Anne K. Dougherty The Healing Power of Chinese Herbs and Medicinal Recipes by Joseph P. Hou and Youyu Jin Alternative Therapies in the Treatment of Brain Injury and Neurobehavioral Disorders: A Practical Guide edited by Gregory J. Murrey Handbook of Cannabis Therapeutics: From Bench to Bedside edited by Ethan B. Russo and Franjo Grotenhermen Handbook of Cannabis Therapeutics From Bench to Bedside Ethan B. Russo, MD Franjo Grotenhermen, MD Editors Routledge Taylor &. Francis Croup NEW YORK AND LONDON First Published by The Haworth Press, Inc., 10 Alice Street, Binghamton, NY 13904-1580. Transferred to Digital Printing 2010 by Routledge 270 Madison Ave, New York NY 10016 2 Park Square, Milton Park, Abingdon, Oxon, OX14 4RN For more information on this book or to order, visit http://www.haworthpress.com/store/product.asp?sku=5741 or call 1-800-HAWORTH (800-429-6784) in the United States and Canada or (607) 722-5857 outside the United States and Canada or contact [email protected] © 2006 by The Haworth Press, Inc. -

INTRODUCTION One of the Tasks of a Pharmacologist Is the Classification of Drugs
NEUROPSYCHOPHARMACOLOGIC STUDIES OF MARIJUANA: SOME SYNTHETIC AND NATURAL THC DERIVATIVES IN ANIMALS AND MAN* Edward F. Domino Department of Pharmacology, University of Michigan, Ann Arbor 48104 and Lafayette Clinic, Detroit, Mich. 48207 INTRODUCTION One of the tasks of a pharmacologist is the classification of drugs. Marijuana and its derivatives pose some problems. Legally marijuana has been classified as a narcotic, yet pharmacologically it is quite different. Mankind has known about the Cannabis plant from which marijuana and other products are obtained for more than 4,708 years. A considerable body of data is available to describe its gross effects. Over the centuries it has been called a depressant, euphoriant, ine- briant, intoxicant, hallucinogen, psychotomimetic, sedative-hypnotic-anesthetic, and stimulant. Perhaps more than any other drug it is a social irritant that elicits rather remarkable behavior reactions in both users and nonusers. Moreau’ in 1845, employed it to produce a model psychosis, thus initiating the fashion to study hallucinogens as a means of gaining insights into mental illness. Lewin2 classified Cannabis as one of the “phantastica: hallucinating substances.” One text on hallucinogens3 does not even refer to Cannabis, but another does? There are some aspects of the subjective effects of both LSD-25 and Cannabis that are similar, yet the latter produces sedative effects, no significant sympatho- mimetic actions, and no cross-tolerance to LSD-25. Hollister5 reviewed the re- cent findings on the effects in man of marijuana and its putative active ingredient Ag-THC, stressing that current investigators have, for the most part, confirmed the Cannabis-induced clinical syndromes long known to occur. -

The Use of Cannabinoids in Animals and Therapeutic Implications for Veterinary Medicine: a Review
Veterinarni Medicina, 61, 2016 (3): 111–122 Review Article doi: 10.17221/8762-VETMED The use of cannabinoids in animals and therapeutic implications for veterinary medicine: a review L. Landa1, A. Sulcova2, P. Gbelec3 1Faculty of Medicine, Masaryk University, Brno, Czech Republic 2Central European Institute of Technology, Masaryk University, Brno, Czech Republic 3Veterinary Hospital and Ambulance AA Vet, Prague, Czech Republic ABSTRACT: Cannabinoids/medical marijuana and their possible therapeutic use have received increased atten- tion in human medicine during the last years. This increased attention is also an issue for veterinarians because particularly companion animal owners now show an increased interest in the use of these compounds in veteri- nary medicine. This review sets out to comprehensively summarise well known facts concerning properties of cannabinoids, their mechanisms of action, role of cannabinoid receptors and their classification. It outlines the main pharmacological effects of cannabinoids in laboratory rodents and it also discusses examples of possible beneficial use in other animal species (ferrets, cats, dogs, monkeys) that have been reported in the scientific lit- erature. Finally, the article deals with the prospective use of cannabinoids in veterinary medicine. We have not intended to review the topic of cannabinoids in an exhaustive manner; rather, our aim was to provide both the scientific community and clinical veterinarians with a brief, concise and understandable overview of the use of cannabinoids in veterinary -

Appendix-2Final.Pdf 663.7 KB
North West ‘Through the Gate Substance Misuse Services’ Drug Testing Project Appendix 2 – Analytical methodologies Overview Urine samples were analysed using three methodologies. The first methodology (General Screen) was designed to cover a wide range of analytes (drugs) and was used for all analytes other than the synthetic cannabinoid receptor agonists (SCRAs). The analyte coverage included a broad range of commonly prescribed drugs including over the counter medications, commonly misused drugs and metabolites of many of the compounds too. This approach provided a very powerful drug screening tool to investigate drug use/misuse before and whilst in prison. The second methodology (SCRA Screen) was specifically designed for SCRAs and targets only those compounds. This was a very sensitive methodology with a method capability of sub 100pg/ml for over 600 SCRAs and their metabolites. Both methodologies utilised full scan high resolution accurate mass LCMS technologies that allowed a non-targeted approach to data acquisition and the ability to retrospectively review data. The non-targeted approach to data acquisition effectively means that the analyte coverage of the data acquisition was unlimited. The only limiting factors were related to the chemical nature of the analyte being looked for. The analyte must extract in the sample preparation process; it must chromatograph and it must ionise under the conditions used by the mass spectrometer interface. The final limiting factor was presence in the data processing database. The subsequent study of negative MDT samples across the North West and London and the South East used a GCMS methodology for anabolic steroids in addition to the General and SCRA screens. -

The Effects of Cannabinoids on Stress-Coping
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 5-2021 The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredtictable Stress: A Preclinical Systematic Review and Meta-Analysis Noa Reuveni Utah State University Follow this and additional works at: https://digitalcommons.usu.edu/etd Part of the Psychology Commons Recommended Citation Reuveni, Noa, "The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredtictable Stress: A Preclinical Systematic Review and Meta-Analysis" (2021). All Graduate Theses and Dissertations. 8098. https://digitalcommons.usu.edu/etd/8098 This Thesis is brought to you for free and open access by the Graduate Studies at DigitalCommons@USU. It has been accepted for inclusion in All Graduate Theses and Dissertations by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. THE EFFECTS OF CANNABINOIDS ON STRESS-COPING BEHAVIORS AND NEUROENDOCRINOLOGICAL MEASURES IN CHRONIC UNPREDTICTABLE STRESS: A PRECLINICAL SYSTEMATIC REVIEW AND META-ANALYSIS by Noa Reuveni A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in Psychology Approved: __________________ __________________ Sara Freeman, Ph.D. Scott Bates, Ph.D. Co-Major Professor Co-Major Professor __________________ __________________ Diana Meter, Ph.D. Sarah Schwartz, Ph.D. Committee Member Committee Member __________________ __________________ Tyler Renshaw, Ph.D. D. Richard Cutler, Ph.D. Committee Member Interim Vice Provost for Graduate Studies UTAH STATE UNIVERSITY Logan, Utah 2021 ii Copyright © Noa Reuveni 2021 All Rights Reserved iii ABSTRACT The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredictable Stress: A Preclinical Systematic Review and Meta-Analysis by Noa Reuveni, Master of Science Utah State University, 2021 Major Professors: Dr. -

Cannabinoid Hope for MND Potent Pot Linked to Psychosis
Highlights from clinical trials on cannabis science RESEARCH ROUND-UP By Liam Drew SPASTICITY Cannabinoid hope for MND A clinical trial of a cannabi- noid oral spray that is used to treat people with multiple sclerosis (MS) suggests that the treatment could also ease the symptoms of motor neuron disease (MND). The treatment targets spasticity, a condition caused by the permanent contraction of muscles, which impedes a person’s movement. It is a prominent symptom of both MND and MS. In 2010, after a long history of people with MS using cannabis to self-medicate, a cannabis- based treatment to manage spasticity associated with MS was approved in the United Kingdom. Giancarlo Comi at Vita-Salute San Raffaele University in Milan, Italy, and his colleagues now suggest that this same cannabinoid preparation can ease spasticity in people with MND. decreased people’s reported psychosis — a condition in of strains containing less than In a double-blind, rand- pain levels. Future studies which a person’s perception of 10% THC. But using strains omized phase II trial, 30 people might more finely distinguish reality is distorted. that contained more than 10% received a placebo and 29 peo- the treatment’s efficacy in two The researchers surveyed THC made the probability of ple were given the cannabinoid forms of MND: amyotrophic 901 people with psychotic developing psychosis almost spray — a solution containing lateral sclerosis, in which spas- disorders, who had been five times higher than that of roughly equal amounts of tet- ticity varies in its prevalence admitted into psychiatric care non-users. -

A Budding Source of Targets for Treating Inflammatory and Neuropathic Pain
Clinical and Translational Science Institute Centers 1-1-2018 The Endogenous Cannabinoid System: A Budding Source of Targets for Treating Inflammatory and Neuropathic Pain Giulia Donvito Virginia Commonwealth University Sara R. Nass West Virginia University Jenny L. Wilkerson Virginia Commonwealth University Zachary A. Curry Virginia Commonwealth University Lesley D. Schurman Virginia Commonwealth University See next page for additional authors Follow this and additional works at: https://researchrepository.wvu.edu/ctsi Part of the Medicine and Health Sciences Commons Digital Commons Citation Donvito, Giulia; Nass, Sara R.; Wilkerson, Jenny L.; Curry, Zachary A.; Schurman, Lesley D.; Kinsey, Steven G.; and Lichtman, Aron H., "The Endogenous Cannabinoid System: A Budding Source of Targets for Treating Inflammatory and Neuropathic Pain" (2018). Clinical and Translational Science Institute. 717. https://researchrepository.wvu.edu/ctsi/717 This Article is brought to you for free and open access by the Centers at The Research Repository @ WVU. It has been accepted for inclusion in Clinical and Translational Science Institute by an authorized administrator of The Research Repository @ WVU. For more information, please contact [email protected]. Authors Giulia Donvito, Sara R. Nass, Jenny L. Wilkerson, Zachary A. Curry, Lesley D. Schurman, Steven G. Kinsey, and Aron H. Lichtman This article is available at The Research Repository @ WVU: https://researchrepository.wvu.edu/ctsi/717 Neuropsychopharmacology REVIEWS (2018) 43, 52–79 © 2018 American -
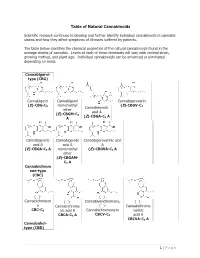
Table of Natural Cannabinoids
Table of Natural Cannabinoids Scientific research continues to develop and further identify individual cannabinoids in cannabis strains and how they affect symptoms of illnesses suffered by patients. The table below identifies the chemical properties of the natural cannabinoids found in the average strains of cannabis. Levels of each of these chemicals will vary with varietal strain, growing method, and plant age. Individual cannabinoids can be enhanced or eliminated depending on need. Cannabigerol- type (CBG) Cannabigerol Cannabigerol Cannabigerovarin (E)-CBG-C monomethyl (E)-CBGV-C 5 Cannabinerolic 3 ether acid A (E)-CBGM-C 5 (Z)-CBGA-C A A 5 Cannabigerolic Cannabigerolic Cannabigerovarinic acid acid A acid A A (E)-CBGA-C5 A monomethyl (E)-CBGVA-C3 A ether (E)-CBGAM- C5 A Cannabichrom ene-type (CBC) (±)- (±)- Cannabichromen (±)- Cannabivarichromene, (±)- e Cannabichrome (±)- Cannabichrome CBC-C5 nic acid A Cannabichromevarin varinic CBCA-C5 A CBCV-C3 acid A CBCVA-C3 A Cannabidiol- type (CBD) 1 | Page (−)-Cannabidiol Cannabidiol Cannabidiol-C4 (−)- Cannabidiorc CBD-C5 momomethyl CBD-C4 Cannabidivarin ol ether CBDV-C3 CBD-C1 CBDM-C5 Cannabidiolic Cannabidivarini acid c acid CBDA-C5 CBDVA-C3 Cannabinodiol- type (CBND) Cannabinodiol Cannabinodivar CBND-C5 in CBND-C3 Tetrahydrocan nabinol-type (THC) 9 9 9 Δ - Δ - Δ - Δ9- Tetrahydrocanna Tetrahydrocan Tetrahydrocannabivarin 9 Tetrahydrocan binol nabinol-C4 Δ -THCV-C3 9 9 nabiorcol Δ -THC-C5 Δ -THC-C4 9 Δ -THCO-C1 9 9 Δ -Tetrahydro- Δ9-Tetrahydro- Δ -Tetrahydro- Δ9-Tetrahydro- cannabinolic -

Cannabinoid Receptor Antagonists Counteract Sensorimotor Gating Deficits in the Phencyclidine Model of Psychosis
Neuropsychopharmacology (2007) 32, 2098–2107 & 2007 Nature Publishing Group All rights reserved 0893-133X/07 $30.00 www.neuropsychopharmacology.org Cannabinoid Receptor Antagonists Counteract Sensorimotor Gating Deficits in the Phencyclidine Model of Psychosis ,1 2,3 4 5 3,6 Martina Ballmaier* , Marco Bortolato , Cristina Rizzetti , Michele Zoli , GianLuigi Gessa , 1,6 4,6 Andreas Heinz and PierFranco Spano 1Department of Psychiatry and Psychotherapy, Charite´ University Medicine, Campus Mitte, Berlin, Germany; 2Department of Pharmacology, 3 4 University of California, Irvine, USA; Department of Neuroscience, Bernard B Brodie, University of Cagliari, Cagliari, Italy; Department of 5 Biomedical Sciences and Biotechnologies, Brescia University Medical School, Brescia, Italy; Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italy Clinical and laboratory findings suggest that cannabinoids and their receptors are implicated in schizophrenia. The role of cannabinoids in schizophrenia remains however poorly understood, as data are often contradictory. The primary aim of this study was to investigate whether the cannabinoid CB1 receptor antagonists rimonabant and AM251 are able to reverse deficits of sensorimotor gating induced by phencyclidine and to mimic the ‘atypical’ antipsychotic profile of clozapine. The prepulse inhibition (PPI) of the startle reflex was used to measure deficits of sensorimotor gating. PPI-disruptive effects of phencyclidine and their antagonism by rimonabant, AM251, and clozapine were studied in rats. The effects of rimonabant were carefully examined taking into account dose ranges, vehicle, and route of administration. We also examined the ability of rimonabant to reduce the PPI-disruptive effects of dizocilpine and apomorphine. Rimonabant as well as AM251 significantly counteracted the phencyclidine-disruptive model of PPI, comparable to the restoring effect of clozapine; no augmentation effect was observed with rimonabant and clozapine as cotreatment. -

Structure-Activity Relationships of the Cannabinoids
RESEARCH ANALYSIS and UTILIZATION SYSTEM Structure-Activity Relationships of the Cannabinoids DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service Alcohol, Drug Abuse, and Mental Health Administration Structure-Activity Relationships of the Cannabinoids Editors: Rao S. Rapaka, Ph.D. Division of Preclinical Research National Institute on Drug Abuse Alexandros Makriyannis, Ph.D. School of Pharmacy and Institute of Materials Science University of Connecticut and F. Bitter National Magnet Laboratory Massachusetts Institute of Technology NIDA Research Monograph 79 A RAUS Review Report 1987 U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service Alcohol, Drug Abuse, and Mental Health Administration National Institute on Drug Abuse 5600 Fishers Lane Rockville, Maryland 20857 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, DC 20402 NIDA Research Monographs are prepared by the research divisions of the National Institute on Drug Abuse and published by its Office of Science. The primary objective of the series is to provide critical reviews of research problem areas and techniques, the content of state-of-the-art conferences, and integrative research reviews. Its dual publication emphasis is rapid and targeted dissemination to the scientific and professional community. Editorial Advisors MARTIN W. ADLER, Ph.D. MARY L. JACOBSON Temple University School of Medicine National Federation of Parents for Philadelphia, Pennsylvania Drug-Free Youth Omaha, Nebraska SYDNEY ARCHER, Ph.D. Rensselaer Polytechnic lnstitute Troy, New York REESE T. JONES, M.D. Langley Porter Neuropsychiatric Institute RICHARD E. BELLEVILLE, Ph.D. San Francisco, California NB Associates. Health Sciences Rockville, Maryland DENISE KANDEL, Ph.D. KARST J. BESTEMAN College of Physicians and Surgeons of Alcohol and Drug Problems Association Columbia University of North America New York, New York Washington, D.C.