Structure-Activity Relationships of the Cannabinoids
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Analysis of Drugs Manual September 2019
Drug Enforcement Administration Office of Forensic Sciences Analysis of Drugs Manual September 2019 Date Posted: 10/23/2019 Analysis of Drugs Manual Revision: 4 Issue Date: September 5, 2019 Effective Date: September 9, 2019 Approved By: Nelson A. Santos Table of Contents CHAPTER 1 – QUALITY ASSURANCE ......................................................................... 3 CHAPTER 2 – EVIDENCE ANALYSIS ......................................................................... 93 CHAPTER 3 – FIELD ASSISTANCE .......................................................................... 165 CHAPTER 4 – FINGERPRINT AND SPECIAL PROGRAMS ..................................... 179 Appendix 1A – Definitions ........................................................................................... 202 Appendix 1B – Acronyms and Abbreviations .............................................................. 211 Appendix 1C – Instrument Maintenance Schedule ..................................................... 218 Appendix 1D – Color Test Reagent Preparation and Procedures ............................... 224 Appendix 1E – Crystal and Precipitate Test Reagent Preparation and Procedures .... 241 Appendix 1F – Thin Layer Chromatography................................................................ 250 Appendix 1G – Qualitative Method Modifications ........................................................ 254 Appendix 1H – Analytical Supplies and Services ........................................................ 256 Appendix 2A – Random Sampling Procedures -

Cannabinoids in the Pathophysiology of Skin Inflammation
molecules Review Cannabinoids in the Pathophysiology of Skin Inflammation Cristian Scheau 1 , Ioana Anca Badarau 1, Livia-Gratiela Mihai 1, Andreea-Elena Scheau 2, Daniel Octavian Costache 3, Carolina Constantin 4,5, Daniela Calina 6 , Constantin Caruntu 1,7,*, Raluca Simona Costache 8,* and Ana Caruntu 9,10 1 Department of Physiology, “Carol Davila” University of Medicine and Pharmacy, 050474 Bucharest, Romania; [email protected] (C.S.); [email protected] (I.A.B.); [email protected] (L.-G.M.) 2 Department of Radiology and Medical Imaging, Fundeni Clinical Institute, 022328 Bucharest, Romania; [email protected] 3 Department of Dermatology, “Carol Davila” Central Military Emergency Hospital, 010825 Bucharest, Romania; [email protected] 4 Immunology Department, ”Victor Babes” National Institute of Pathology, 050096 Bucharest, Romania; [email protected] 5 Department of Pathology, Colentina University Hospital, 020125 Bucharest, Romania 6 Department of Clinical Pharmacy, University of Medicine and Pharmacy of Craiova, 200349 Craiova, Romania; [email protected] 7 Department of Dermatology, “Prof. N. Paulescu” National Institute of Diabetes, Nutrition and Metabolic Diseases, 011233 Bucharest, Romania 8 Gastroenterology and Internal Medicine Clinic, Carol Davila University Central Emergency Military Hospital, Carol Davila University of Medicine and Pharmacy, 050474 Bucharest, Romania 9 Department of Oral and Maxillofacial Surgery, “Carol Davila” Central Military Emergency Hospital, 010825 Bucharest, Romania; [email protected] 10 Faculty of Medicine, “Titu Maiorescu” University, 031593 Bucharest, Romania * Correspondence: [email protected] (C.C.); [email protected] (R.S.C.); Tel.: +40-745-086-978 (C.C.) Academic Editor: Eric J. Downer Received: 30 December 2019; Accepted: 2 February 2020; Published: 4 February 2020 Abstract: Cannabinoids are increasingly-used substances in the treatment of chronic pain, some neuropsychiatric disorders and more recently, skin disorders with an inflammatory component. -

Cannabigerol Is a Potential Therapeutic Agent in a Novel Combined Therapy for Glioblastoma
cells Article Cannabigerol Is a Potential Therapeutic Agent in a Novel Combined Therapy for Glioblastoma Tamara T. Lah 1,2,3,*, Metka Novak 1, Milagros A. Pena Almidon 4, Oliviero Marinelli 4 , Barbara Žvar Baškoviˇc 1, Bernarda Majc 1,3, Mateja Mlinar 1, Roman Bošnjak 5, Barbara Breznik 1 , Roby Zomer 6 and Massimo Nabissi 4 1 Department of Genetic Toxicology and Cancer Biology, National Institute of Biology, 1000 Ljubljana, Slovenia; [email protected] (M.N.); [email protected] (B.Ž.B.); [email protected] (B.M.); [email protected] (M.M.); [email protected] (B.B.) 2 Faculty of Chemistry and Chemical Technology, University of Ljubljana, 1000 Ljubljana, Slovenia 3 Jožef Stefan International Postgraduate School, 1000 Ljubljana, Slovenia 4 School of Pharmacy, Experimental Medicine Section, University of Camerino, 62032 Camerino, Italy; [email protected] (M.A.P.A.); [email protected] (O.M.); [email protected] (M.N.) 5 Department of Neurosurgery, University Medical Centre Ljubljana, 1000 Ljubljana, Slovenia; [email protected] 6 MGC Pharmaceuticals d.o.o., 1000 Ljubljana, Slovenia; [email protected] * Correspondence: [email protected]; Tel.: +386-41-651-629 Simple Summary: Among primary brain tumours, glioblastoma is the most aggressive. As early relapses are unavoidable despite standard-of-care treatment, the cannabinoids delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) alone or in combination have been suggested as a combined treatment strategy for glioblastomas. However, the known psychoactive effects of THC hamper its medical applications in these patients with potential cognitive impairment due to the progression of the Citation: Lah, T.T.; Novak, M.; Pena Almidon, M.A.; Marinelli, O.; disease. -

Nabilone for Chronic Pain Management: a Review of Clinical Effectiveness, Safety, and Guidelines
TITLE: Nabilone for Chronic Pain Management: A Review of Clinical Effectiveness, Safety, and Guidelines DATE: 11 November 2011 CONTEXT AND POLICY ISSUES Cannabis sativa is a flowering plant that has long been used as a recreational drug, and for medicinal purposes.1,2 The main psychoactive component of cannabis is delta-9- tetrahydrocannabinol (∆9-THC).2 Nabilone (Cesamet®) is an oral synthetic cannabinoid, which is licensed in Canada for treating patients with severe nausea and vomiting related to chemotherapy for cancer and who have failed to respond adequately to conventional antiemetic treatments.2-4 Clinical trials and anecdotal reports have suggested that the use of nabilone in other medical conditions, such as appetite stimulation, anxiety, spasticity, and pain.1,5 Chronic pain affects approximately one in five people in developed countries and two in five in less well-resourced countries. In many circumstances, the patient’s quality of life is poor due to persistent pain caused either by an ongoing illness or nerve damage caused by the disease after resolution or cure of the disease.6 Multiple sclerosis (MS) is a neurodegenerative disease, and is the most common cause of neurological disability in young people, with an average age of onset around 30 years and a prevalence of about 120 per 100,000 individuals in North America. The majority of patients with MS display symptoms, such as fatigue, muscle stiffness or spasticity, pain, memory problems, balance trouble, tremors, urinary disturbance, and sexual dysfunctions.2,7 The purpose of this review is to assess the evidence of benefits and harms related to the use of nabilone in management of chronic pain, including patients with MS. -

A Dissertation Entitled Uncovering Cannabinoid Signaling in C. Elegans
A Dissertation Entitled Uncovering Cannabinoid Signaling in C. elegans: A New Platform to Study the Effects of Medicinal Cannabis By Mitchell Duane Oakes Submitted to the Graduate Faculty as partial fulfillment of the requirements for the Doctor of Philosophy Degree in Biology ________________________________________ Dr. Richard Komuniecki, Committee Chair _______________________________________ Dr. Bruce Bamber, Committee Member ________________________________________ Dr. Patricia Komuniecki, Committee Member ________________________________________ Dr. Robert Steven, Committee Member ________________________________________ Dr. Ajith Karunarathne, Committee Member ________________________________________ Dr. Jianyang Du, Committee Member ________________________________________ Dr. Amanda Bryant-Friedrich, Dean College of Graduate Studies The University of Toledo August 2018 Copyright 2018, Mitchell Duane Oakes This document is copyrighted material. Under copyright law, no parts of this document may be reproduced without the expressed permission of the author. An Abstract of Uncovering Cannabinoid Signaling in C. elegans: A New Platform to Study the Effects of Medical Cannabis By Mitchell Duane Oakes Submitted to the Graduate Faculty as partial fulfillment of the requirements for the Doctor of Philosophy Degree in Biology The University of Toledo August 2018 Cannabis or marijuana, a popular recreational drug, alters sensory perception and exerts a range of medicinal benefits. The present study demonstrates that C. elegans exposed to -

Safe Access to Medical Cannabis in New Mexico Schools a Guide Prepared for Tisha Brick by Jason Barker with Safe Access New Mexico
Safe Access To Medical Cannabis in New Mexico Schools A guide prepared for Tisha Brick by Jason Barker with Safe Access New Mexico Safe Access New Mexico ~ A Chapter of Americans For Safe Access UNITE-NETWORK-GROW-INFORM-KNOW-EDUCATE-ACTIVISM-VOTE-HEALTH-WELLNESS (All Rights Reserved 04/20/2018) 1 Program Participants Should Be Able To Use Medical Cannabis At Schools We've come a long way since cannabis was first decriminalized in Oregon in 1973 and then in New Mexico; medical cannabis history started in 1978, after public hearings the legislature enacted H.B. 329, the nation’s first law recognizing the medical value of cannabis…the first law. And it has now been over a decade since the passage of the Lynn and Erin Compassionate Use Act. Safe Access for those patients who will benefit most from medical cannabis treatments; still need to overcome political, social and legal barriers with advocacy by creating policies that improve access to medical cannabis for patients - and that means at school too. Schools already allow children to use all kinds of psychotropic medications—from Ritalin to opioid painkillers—when prescribed by a physician. 2 States That Allow Safe Access To Medical Cannabis In Schools New Jersey - 2015 * Maine - 2015 * Colorado - 2016 (1st Jack’s Law) & 2018 Washington - 2016 Pennsylvania - 2017 Illinois - 2018 (* States Program is Modelled after New Mexico’s Medical Cannabis Program) 3 New Jersey in November 2015 became the first state to do so. Governor Christie signed a bill directing all school districts in New Jersey to adopt rules that permit children with developmental disabilities to consume cannabis oil or another edible cannabis product. -

Problems of Drug Dependence 1980 Proceedings of the 42Nd Annual Scientific Meeting the Committee on Problems of Drug Dependence
National Institute on Drug Abuse MONOGRAPH SERIES Problems of Drug Dependence 1980 Proceedings of the 42nd Annual Scientific Meeting The Committee on Problems of Drug Dependence, Inc. U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES • Public Health Service • Alcohol, Drug Abuse, and Mental Health Administration Problems of Drug Dependence, 1980 Proceedings of the 42nd Annual Scientific Meeting, The Committee on Problems of Drug Dependence, Inc. Editor: Louis S. Harris, Ph.D. NIDA Research Monograph 34 February 1981 DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service Alcohol, Drug Abuse, and Mental Health Administration National Institute on Drug Abuse Division of Research 5600 Fishers Lane Rockville, Maryland 20857 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 The NIDA Research Monograph series is prepared by the Division of Research of the National Institute on Drug Abuse. Its primary objective is to provide critical reviews of research problem areas and techniques, the content of state-of-the-art conferences, integrative research reviews and significant original research. Its dual publication emphasis is rapid and targeted dissemination to the scientific and professional community. Editorial Advisory Board Avram Goldstein, M.D. Addiction Research Foundation Palo Alto, California Jerome Jaffe, M.D. College of Physicians and Surgeons Columbia University, New York Reese T. Jones, M.D. Langley Porter Neuropsychiatric Institute University of California San Francisco, California William McGlothlin, Ph.D. Deportment of Psychology, UCLA Los Angeles, California Jack Mendelson, M.D. Alcohol and Drug Abuse Research Center Harvard Medical School McLean Hospital Belmont, Massachusetts Helen Nowlis, Ph.D. Office of Drug Education, DHHS Washington, D.C Lee Robins, Ph.D. -
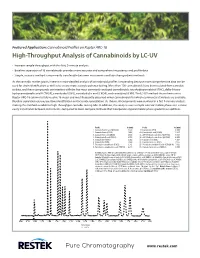
High-Throughput Analysis of Cannabinoids by LC-UV
Featured Application: Cannabinoid Profiles on Raptor ARC-18 High-Throughput Analysis of Cannabinoids by LC-UV • Increase sample throughput with this fast, 9-minute analysis. • Baseline separation of 16 cannabinoids provides more accurate and comprehensive potency and profile data. • Simple, isocratic method is more easily transferable between instruments and labs than gradient methods. As the cannabis market grows, interest in more detailed analysis of cannabinoid profiles is expanding because more comprehensive data can be used for strain identification as well as to ensure more accurate potency testing. More than 100 cannabinoids have been isolated from cannabis to date, and these compounds can interfere with the five most commonly analyzed cannabinoids: tetrahydrocannabinol (THC), delta-9-tetra- hydrocannabinolic acid A (THCA), cannabidiol (CBD), cannabidiolic acid (CBDA), and cannabinol (CBN). The LC-UV method shown here uses a Raptor ARC-18 column to fully resolve 16 major and most frequently observed minor cannabinoids for which commercial standards are available. Baseline separation ensures positive identification and accurate quantitation. As shown, all compounds were resolved in a fast 9-minute analysis, making this method suitable for high-throughput cannabis testing labs. In addition, this analysis uses a simple isocratic mobile phase so it is more easily transferable between instruments, compared to more complex methods that incorporate atypical mobile phase gradients or additives. Peaks tR (min) Peaks tR (min) 1. Cannabidivarinic acid (CBDVA) 1.877 9. Cannabinol (CBN) 4.609 2. Cannabidivarin (CBDV) 2.086 10. Cannabinolic acid (CBNA) 5.437 3. Cannabidiolic acid (CBDA) 2.592 11. Δ9-Tetrahydrocannabinol (Δ9-THC) 5.815 4. Cannabigerolic acid (CBGA) 2.750 12. -

Effect of Curcumin, Mixture of Curcumin and Piperine and Curcum (Turmeric) on Lipid Profile of Normal and Hyperlipidemic Rats
The Egyptian Journal of Hospital Medicine Vol., 21: 145 – 161 December 2005 I.S.S.N: 12084 2002–1687 Effect of Curcumin, Mixture of Curcumin and Piperine and Curcum (Turmeric) on Lipid Profile of Normal and Hyperlipidemic Rats GHADA, Z. A. Soliman Lecturer of Biochemistry, Biochemistry Department, National Nutrition Institute, Cairo Abstract Curcumin is a polyphenolic, yellow pigment obtained from rhizomes of Curcuma longa (curcum), used as a spice and food colouring. The extracts have several pharmacological effects. We evaluated the effect of curcum, curcumin, and mixture of curcumin and piperine on plasma lipids in normal and hypercholesterolemic rats. A total of 270 rats, divided into 27 groups, were used. G1, G11: control, G2-G11: normal rats fed control diet supplemented with different levels of curcumin and curcum (G2-G6: 0.1%, 0.25%, 0.5%, 1.0%, 2.0% respectively, G7-G11: 1.67%, 4.167%, 8.34%, 16.67%, and 33.34). G12-G26: at first fed control diet supplemented with 2% cholesterol then G13-17, 21-25 fed a control diet supplemented with different levels of curcumin, and curcum [the same levels as G2-G11; G18-20 fed control diet supplemented with mixture of curcumin (0.1, 0.25, 0.5%) and piperine (20 mg/kg BW)], G12 was sacrificed before addition of studied materials, G26 were fed control diet. Lipid profile, triacylglycerol and phospholipids of plasma and organs as liver and heart were measured. Serum cholesterol (total, LDL-C, VLDL-C), triacylglycerol and phospholipids contents were elevated in cholesterol-fed rats, while HDL-C were decreased. -

Mechanistic Study of Physicochemical and Biochemical Processes
Mechanistic study of physicochemical and biochemical processes affecting intestinal absorption of the sesquiterpene lactone nobilin from multi-component systems in the Caco-2 model. Inauguraldissertation zur Erlangung der Würde eines Doktors der Philosophie vorgelegt der Philosophisch-Naturwissenschaftlichen Fakultät der Universität Basel von URSULA STEPHANIE THORMANN aus Bern (BE) Basel, 2015 Originaldokument gespeichert auf dem Dokumentenserver der Universität Basel edoc.unibas.ch Dieses Werk ist unter dem Vertrag „Creative Commons Namensnennung-Keine kommerzielle Nutzung-Keine Bearbeitung 3.0 Schweiz“ (CC BY-NC-ND 3.0 CH) lizenziert. Die vollständige Lizenz kann unter creativecommons.org/licenses/by-nc-nd/3.0/ch/eingesehen werden. Genehmigt von der Philosophisch-Naturwissenschaftlichen Fakultät auf Antrag von Prof. Dr. G. Imanidis und Prof. Dr. H. E. U. Meyer zu Schwabedissen Basel, den 18. Februar 2014 Prof. Dr. J. Schibler Namensnennung-Keine kommerzielle Nutzung-Keine Bearbeitung 3.0 Schweiz (CC BY-NC-ND 3.0 CH) Sie dürfen: Teilen — den Inhalt kopieren, verbreiten und zugänglich machen Unter den folgenden Bedingungen: Namensnennung — Sie müssen den Namen des Autors/Rechteinhabers in der von ihm festgelegten Weise nennen. Keine kommerzielle Nutzung — Sie dürfen diesen Inhalt nicht für kommerzielle Zwecke nutzen. Keine Bearbeitung erlaubt — Sie dürfen diesen Inhalt nicht bearbeiten, abwandeln oder in anderer Weise verändern. Wobei gilt: Verzichtserklärung — Jede der vorgenannten Bedingungen kann aufgehoben werden, sofern Sie die -

Medicinal Chemistry Endeavors Around the Phytocannabinoids
CHEMISTRY & BIODIVERSITY – Vol. 4 (2007) 1707 REVIEW Medicinal Chemistry Endeavors around the Phytocannabinoids by Eric Stern and Didier M. Lambert* Drug Design and Discovery Center and Unite´ de Chimie pharmaceutique et de Radiopharmacie, Ecole de Pharmacie, Faculte´ de Me´decine, Universite´ catholique de Louvain, Avenue E. Mounier 73, U.C.L. 73.40, B-1200 Bruxelles (phone: þ3227647347; fax: þ3227647363; e-mail: [email protected]) Over the past 50 years, a considerable research in medicinal chemistry has been carried out around the natural constituents of Cannabis sativa L. Following the identification of D9-tetrahydrocannabinol (D9-THC) in 1964, critical chemical modifications, e.g., variation of the side chain at C(3) and the opening of the tricyclic scaffold, have led to the characterization of potent and cannabinoid receptor subtype-selective ligands. Those ligands that demonstrate high affinity for the cannabinoid receptors and good biological efficacy are still used as powerful pharmacological tools. This review summarizes past as well as recent developments in the structure–activity relationships of phytocannabinoids. 1. Introduction. – Despite the wide uses of preparations of the hemp Cannabis sativa L. during the History, the modern pharmacology of natural cannabinoids has been hampered by the slow progress in the elucidations of the chemical structures of its major components. Indeed, it is nowadays known that more than 70 compounds derived from a diterpene structure are present in the plant [1], and this fact may explain the difficulty to obtain pure chemical entities in the past. In addition, the medicinal research for more than a half century has been driven by the search for the components responsible for the psychoactive effects of cannabis, this era in the history of the chemical research on cannabinoids have been recently reviewed [2][3]. -
Marinol Cannabidiol C21H30O2 Trade Name
Cannabinoids are a group of terpenophenolic compounds secreted by Cannabis flowers that provide relief from a wide array of symptoms including, pain, nausea, and inflammation. They operate by imitating the body’s natural endocannabinoids, which activate to maintain internal stability and overall health. When consumed, cannabinoids bind to receptor sites throughout the brain (CB1 receptors) and body (CB2 receptors). Different cannabinoids have different effects based on their binding affinity for each receptor. By targeting specific cannabinoids at these receptors, different types of relief can be achieved. Presently, there are at least 113 different cannabinoids isolated from Cannabis—each exhibiting varied effects. THC Tetrahydrocannabinol C21H30O2 Trade name: Marinol Legal Status: US – Schedule I, Schedule II (as Cesamet), Schedule III (as Marinol) OH CA – Schedule II UK – Class B AU – S8 (controlled) H Psychoactive Tetrahydrocannabinol (THC) is typically the most abundant cannabinoid present in cannabis products on the market today. THC has very high psychoactive characteristics and is associated with the ‘high’ and euphoria experienced when using cannabis products. When smoked or ingested, THC binds to cannabinoid receptors throughout the body and affects memory, O coordination, concentration, pleasure, and time perception. H Medicinal Benefits Analgesic • Anti-nauseant • Appetite Stimulant Reduces Glaucoma Symptoms Sleep Aid • Reduces Anxiety and PTSD Symptoms CBD Cannabidiol C21H30O2 Trade name: Epidiolex OH Legal Status: US – Schedule I CA – Schedule II UK – POM (Perscription only) H AU – S4 (Perscription only) non-psychoactive Cannabidiol (CBD) is a major phytocannabinoid and accounts for up to 40% of the plant’s extract. Due to its lack of psychoactivity and HO non-interference with motor and psychological functions, it is a leading candidate for a wide variety of medical applications.