INTRODUCTION One of the Tasks of a Pharmacologist Is the Classification of Drugs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cannabinoids in the Pathophysiology of Skin Inflammation
molecules Review Cannabinoids in the Pathophysiology of Skin Inflammation Cristian Scheau 1 , Ioana Anca Badarau 1, Livia-Gratiela Mihai 1, Andreea-Elena Scheau 2, Daniel Octavian Costache 3, Carolina Constantin 4,5, Daniela Calina 6 , Constantin Caruntu 1,7,*, Raluca Simona Costache 8,* and Ana Caruntu 9,10 1 Department of Physiology, “Carol Davila” University of Medicine and Pharmacy, 050474 Bucharest, Romania; [email protected] (C.S.); [email protected] (I.A.B.); [email protected] (L.-G.M.) 2 Department of Radiology and Medical Imaging, Fundeni Clinical Institute, 022328 Bucharest, Romania; [email protected] 3 Department of Dermatology, “Carol Davila” Central Military Emergency Hospital, 010825 Bucharest, Romania; [email protected] 4 Immunology Department, ”Victor Babes” National Institute of Pathology, 050096 Bucharest, Romania; [email protected] 5 Department of Pathology, Colentina University Hospital, 020125 Bucharest, Romania 6 Department of Clinical Pharmacy, University of Medicine and Pharmacy of Craiova, 200349 Craiova, Romania; [email protected] 7 Department of Dermatology, “Prof. N. Paulescu” National Institute of Diabetes, Nutrition and Metabolic Diseases, 011233 Bucharest, Romania 8 Gastroenterology and Internal Medicine Clinic, Carol Davila University Central Emergency Military Hospital, Carol Davila University of Medicine and Pharmacy, 050474 Bucharest, Romania 9 Department of Oral and Maxillofacial Surgery, “Carol Davila” Central Military Emergency Hospital, 010825 Bucharest, Romania; [email protected] 10 Faculty of Medicine, “Titu Maiorescu” University, 031593 Bucharest, Romania * Correspondence: [email protected] (C.C.); [email protected] (R.S.C.); Tel.: +40-745-086-978 (C.C.) Academic Editor: Eric J. Downer Received: 30 December 2019; Accepted: 2 February 2020; Published: 4 February 2020 Abstract: Cannabinoids are increasingly-used substances in the treatment of chronic pain, some neuropsychiatric disorders and more recently, skin disorders with an inflammatory component. -

Downloaded for Personal Non-Commercial Research Or Study, Without Prior Permission Or Charge
https://theses.gla.ac.uk/ Theses Digitisation: https://www.gla.ac.uk/myglasgow/research/enlighten/theses/digitisation/ This is a digitised version of the original print thesis. Copyright and moral rights for this work are retained by the author A copy can be downloaded for personal non-commercial research or study, without prior permission or charge This work cannot be reproduced or quoted extensively from without first obtaining permission in writing from the author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given Enlighten: Theses https://theses.gla.ac.uk/ [email protected] STUDIES OU THE MODE OF ACTION OF QUATERNARY AMMONIUM COMPOUNDS WITH MUSCLE RELAXANT AND OTHER PHARMACOLOGICAL ACTIVITIES A Thesis submitted to the University of Glasgow in candidature for the degree of Doctor of Philosophy in the Faculty of Medicine *y Thomas C. Muir, B.Sc., M.P.S. Division of Experimental Pharmacology, Institute of Physiology, The University, Glasgow. March 1962. ProQuest Number: 10656287 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. uest ProQuest 10656287 Published by ProQuest LLC(2017). Copyright of the Dissertation is held by the Author. -

B Ramamohima R
I I CYTOPROTECTIVE ACTIONS PF NICOTINE: THE INCREASED EXPRESSION OF a.7 NICOTINIC RECEPTORS AND NGF/TrkA RECEPTORS ' . ' f !'b y' Ramamohima R. Jonnala ' ' ' I' Submitted to the Faculty o~the School of Graduate Studies of Medical College of Georgia in Partial Fulfillment of the Requirements of the Degree of Doctor of Philosophy Juiy, 2001 I' l Cytoprotective Actions of Nicotine: The Increased Expression of a.7 Nicotinic Receptors a-!ld NGF/TrkA Receptors This dissertation is submitted by Ramamohana R. Jonnala and has been examined and approved by an appointed committee of the faculty of the school of Graduate Studies of the Medical College of Georgia. The signatures which appear below verify the fact that all required changes have been incorporated and that the dissertation has received final approval with reference to content, form and accuracy of presentation. ' This dissertation is therefore in partial fulfillment of the requirements of the degree of Doctor of Philosophy. ACKNOWLEDGMENTS I would like to extend my appreciation to my major advisor, Dr. Jerry J. Buccafusco, for his guidance, encouragement and support. I would also like to thank my committee members, Dr. Alvin V. Terry Jr., Dr. William D. Hill, Dr. Nevin A. Lambert and Dr. Clare M. Bergson and also my thesis readers Dr. Debra Gearhart and Dr. Dale W. Sickles for their time and valuable suggestions. My sincere thanks to Dr. Deborah L. Lewis, Dr. Robert W. Caldwell and Dr. Gary C. Bond for encouraging me to enter the graduate program in the Department of Pharmacology & Toxicology, Medical College of Georgia. My sincere thanks to members of my laboratory Laura, Vanessa, Daniel, Cat, Mark, Nandu, Lu, and Shyamala for their assistance and suggestions. -
Marinol Cannabidiol C21H30O2 Trade Name
Cannabinoids are a group of terpenophenolic compounds secreted by Cannabis flowers that provide relief from a wide array of symptoms including, pain, nausea, and inflammation. They operate by imitating the body’s natural endocannabinoids, which activate to maintain internal stability and overall health. When consumed, cannabinoids bind to receptor sites throughout the brain (CB1 receptors) and body (CB2 receptors). Different cannabinoids have different effects based on their binding affinity for each receptor. By targeting specific cannabinoids at these receptors, different types of relief can be achieved. Presently, there are at least 113 different cannabinoids isolated from Cannabis—each exhibiting varied effects. THC Tetrahydrocannabinol C21H30O2 Trade name: Marinol Legal Status: US – Schedule I, Schedule II (as Cesamet), Schedule III (as Marinol) OH CA – Schedule II UK – Class B AU – S8 (controlled) H Psychoactive Tetrahydrocannabinol (THC) is typically the most abundant cannabinoid present in cannabis products on the market today. THC has very high psychoactive characteristics and is associated with the ‘high’ and euphoria experienced when using cannabis products. When smoked or ingested, THC binds to cannabinoid receptors throughout the body and affects memory, O coordination, concentration, pleasure, and time perception. H Medicinal Benefits Analgesic • Anti-nauseant • Appetite Stimulant Reduces Glaucoma Symptoms Sleep Aid • Reduces Anxiety and PTSD Symptoms CBD Cannabidiol C21H30O2 Trade name: Epidiolex OH Legal Status: US – Schedule I CA – Schedule II UK – POM (Perscription only) H AU – S4 (Perscription only) non-psychoactive Cannabidiol (CBD) is a major phytocannabinoid and accounts for up to 40% of the plant’s extract. Due to its lack of psychoactivity and HO non-interference with motor and psychological functions, it is a leading candidate for a wide variety of medical applications. -

Recent Developments in Cannabis Chemistry
Recent Developments in Cannabis Chemistry BY ALEXANDER T. SHULGIN, Ph.D. The marijuana plant Cannabis sativa contains a bewildering Introduction array of organic chemicals. As is true with other botanic species, there are representatives of almost all chemical classes present, including mono- and sesquiterpenes, carbohy- drates, aromatics, and a variety of nitrogenous compounds. Interest in the study of this plant has centered primarily on the resinous fraction, as it is this material that is invested with the pharmacological activity that is peculiar to the plant. This resin is secreted by the female plant as a protective agent during seed ripening, although it can be found as a microscopic exudate through the aerial portions of plants of either sex. The pure resin, hashish or charas, is the most potent fraction of the plant, and has served as the source material for most of the chemical studies. The family of chemicals that has been isolated from this source has been referred to as the cannabinoid group. It is unique amongst psychotropic materials from plants in that there are no alkaloids present. The fraction is totally nitro- gen-free. Rather, the set of compounds can be considered as analogs of the parent compound cannabinol (I), a fusion product of terpene and a substituted resorcinol. Beyond the scope of this present review are such questions as the distribution of these compounds within the plant, the bo- tanic variability resulting from geographic distribution, the diversity of pharmacological action assignable to the several Reprinted from Journal of Psychedelic Drugs, vol. II, no. 1, 197 1. 397 398 Marijuana: Medical Papers distinct compounds present, and the various preparations and customs of administration. -

Journal of Pharmacy and Pharmacology 1962 Volume.14 Suppl
BRITISH PHARMACEUTICAL CONFERENCE NINETY-NINTH ANNUAL MEETING, LIVERPOOL, 1962 REPORT OF PROCEEDINGS OFFICERS: President: Miss M. A. Bu rr, M.P.S., Nottingham Chairman: J. C. H anbury, M.A., B.Pharm., F.P.S., F.R.I.C. Vice-Chairmen: R. R . Bennett, B.Sc., F.P.S., F.R.I.C., Eastbourne. H . D eane, B.Sc., F.P.S., F.R.I.C., Sudbury. H . H umphreys Jones, F.P.S., F.R.I.C., Liverpool. T. E. W allis, D .S c., F.P.S., F.R.I.C., F.L.S., London. H . Brindle, M .Sc., F.P.S., F.R.I.C., Altrincham. N. Evers, B.Sc., Ph.D., F.R.I.C., Ware. A. D . P ow ell, M.P.S., F.R.I.C., Nottingham. H. Berry, B.Sc., Dip.Bact. (Lond.), F.P.S., F.R.I.C., Eastbourne. H . B. M ackie, B.Pharm., F.P.S., Brighton. G. R. Boyes, L.M.S.S.A., B.Sc., F.P.S., F.R.I.C., London. H . D avis, C.B.E., B.Sc., Ph.D., F.P.S., F.R.I.C., London. J. P. T o dd , Ph.D., F.P.S., F.R.I.C., Glasgow. K. Bullock, M .Sc., Ph.D., F.P.S., F.R.I.C., Manchester. F. H artley, B.Sc., Ph.D., F.P.S., F.R.I.C., London. G. E. F oster, B.Sc., Ph.D., F.R.I.C., Dartford. H . T reves Bro w n , B.Sc., F.P.S., London. -

The Use of Cannabinoids in Animals and Therapeutic Implications for Veterinary Medicine: a Review
Veterinarni Medicina, 61, 2016 (3): 111–122 Review Article doi: 10.17221/8762-VETMED The use of cannabinoids in animals and therapeutic implications for veterinary medicine: a review L. Landa1, A. Sulcova2, P. Gbelec3 1Faculty of Medicine, Masaryk University, Brno, Czech Republic 2Central European Institute of Technology, Masaryk University, Brno, Czech Republic 3Veterinary Hospital and Ambulance AA Vet, Prague, Czech Republic ABSTRACT: Cannabinoids/medical marijuana and their possible therapeutic use have received increased atten- tion in human medicine during the last years. This increased attention is also an issue for veterinarians because particularly companion animal owners now show an increased interest in the use of these compounds in veteri- nary medicine. This review sets out to comprehensively summarise well known facts concerning properties of cannabinoids, their mechanisms of action, role of cannabinoid receptors and their classification. It outlines the main pharmacological effects of cannabinoids in laboratory rodents and it also discusses examples of possible beneficial use in other animal species (ferrets, cats, dogs, monkeys) that have been reported in the scientific lit- erature. Finally, the article deals with the prospective use of cannabinoids in veterinary medicine. We have not intended to review the topic of cannabinoids in an exhaustive manner; rather, our aim was to provide both the scientific community and clinical veterinarians with a brief, concise and understandable overview of the use of cannabinoids in veterinary -

( 12 ) United States Patent
US010131766B2 (12 ) United States Patent (10 ) Patent No. : US 10 , 131, 766 B2 Hazen et al. (45 ) Date of Patent: Nov . 20 , 2018 ( 54 ) UNSATURATED POLYESTER RESIN 6 ,619 ,886 B1 9 /2003 Harrington 6 ,692 , 802 B1 2 / 2004 Nava SYSTEM FOR CURED IN - PLACE PIPING 7 , 135 ,087 B2 11/ 2006 Blackmore et al. 7 ,799 ,228 B2 9 /2010 Bomak et al . (71 ) Applicant : Interplastic Corporation , Saint Paul, 8 , 047, 238 B2 11/ 2011 Wiessner et al . MN (US ) 8 ,053 ,031 B2 11/ 2011 Stanley et al. 8 ,092 ,689 B2 1 / 2012 Gosselin (72 ) Inventors : Benjamin R . Hazen , Roseville , MN 8 , 298 , 360 B2 10 / 2012 Da Silveira et al. 8 ,418 , 728 B1 4 / 2013 Kiest , Jr . (US ) ; David J . Herzog , Maple Grove , 8 ,586 ,653 B2 11 / 2013 Klopsch et al. MN (US ) ; Louis R . Ross, Cincinnati , 8 ,636 , 869 B2 1 / 2014 Wiessner et al . OH (US ) ; Joel R . Weber , Moundsview , 8 ,741 , 988 B2 6 /2014 Klopsch et al. MN (US ) 8 , 877 , 837 B2 11/ 2014 Yu et al. 9 ,068 , 045 B2 6 /2015 Nava et al. 9 ,074 , 040 B2 7 / 2015 Turshani et al. ( 73 ) Assignee : Interplastic Corporation , St. Paul , MN 9 , 150 , 709 B2 10 / 2015 Klopsch et al . (US ) 9 , 207 , 155 B1 12 / 2015 Allouche et al. 9 ,273 ,815 B2 3 / 2016 Gillanders et al. ( * ) Notice : Subject to any disclaimer, the term of this 9 , 371, 950 B2 6 / 2016 Hairston et al. patent is extended or adjusted under 35 9 , 550 , 933 B2 1 /2017 Chatterji et al . -

The Effects of Cannabinoids on Stress-Coping
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 5-2021 The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredtictable Stress: A Preclinical Systematic Review and Meta-Analysis Noa Reuveni Utah State University Follow this and additional works at: https://digitalcommons.usu.edu/etd Part of the Psychology Commons Recommended Citation Reuveni, Noa, "The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredtictable Stress: A Preclinical Systematic Review and Meta-Analysis" (2021). All Graduate Theses and Dissertations. 8098. https://digitalcommons.usu.edu/etd/8098 This Thesis is brought to you for free and open access by the Graduate Studies at DigitalCommons@USU. It has been accepted for inclusion in All Graduate Theses and Dissertations by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. THE EFFECTS OF CANNABINOIDS ON STRESS-COPING BEHAVIORS AND NEUROENDOCRINOLOGICAL MEASURES IN CHRONIC UNPREDTICTABLE STRESS: A PRECLINICAL SYSTEMATIC REVIEW AND META-ANALYSIS by Noa Reuveni A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in Psychology Approved: __________________ __________________ Sara Freeman, Ph.D. Scott Bates, Ph.D. Co-Major Professor Co-Major Professor __________________ __________________ Diana Meter, Ph.D. Sarah Schwartz, Ph.D. Committee Member Committee Member __________________ __________________ Tyler Renshaw, Ph.D. D. Richard Cutler, Ph.D. Committee Member Interim Vice Provost for Graduate Studies UTAH STATE UNIVERSITY Logan, Utah 2021 ii Copyright © Noa Reuveni 2021 All Rights Reserved iii ABSTRACT The Effects of Cannabinoids on Stress-Coping Behaviors and Neuroendocrinological Measures in Chronic Unpredictable Stress: A Preclinical Systematic Review and Meta-Analysis by Noa Reuveni, Master of Science Utah State University, 2021 Major Professors: Dr. -
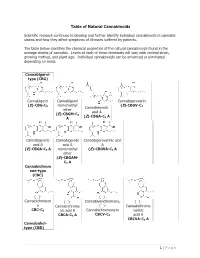
Table of Natural Cannabinoids
Table of Natural Cannabinoids Scientific research continues to develop and further identify individual cannabinoids in cannabis strains and how they affect symptoms of illnesses suffered by patients. The table below identifies the chemical properties of the natural cannabinoids found in the average strains of cannabis. Levels of each of these chemicals will vary with varietal strain, growing method, and plant age. Individual cannabinoids can be enhanced or eliminated depending on need. Cannabigerol- type (CBG) Cannabigerol Cannabigerol Cannabigerovarin (E)-CBG-C monomethyl (E)-CBGV-C 5 Cannabinerolic 3 ether acid A (E)-CBGM-C 5 (Z)-CBGA-C A A 5 Cannabigerolic Cannabigerolic Cannabigerovarinic acid acid A acid A A (E)-CBGA-C5 A monomethyl (E)-CBGVA-C3 A ether (E)-CBGAM- C5 A Cannabichrom ene-type (CBC) (±)- (±)- Cannabichromen (±)- Cannabivarichromene, (±)- e Cannabichrome (±)- Cannabichrome CBC-C5 nic acid A Cannabichromevarin varinic CBCA-C5 A CBCV-C3 acid A CBCVA-C3 A Cannabidiol- type (CBD) 1 | Page (−)-Cannabidiol Cannabidiol Cannabidiol-C4 (−)- Cannabidiorc CBD-C5 momomethyl CBD-C4 Cannabidivarin ol ether CBDV-C3 CBD-C1 CBDM-C5 Cannabidiolic Cannabidivarini acid c acid CBDA-C5 CBDVA-C3 Cannabinodiol- type (CBND) Cannabinodiol Cannabinodivar CBND-C5 in CBND-C3 Tetrahydrocan nabinol-type (THC) 9 9 9 Δ - Δ - Δ - Δ9- Tetrahydrocanna Tetrahydrocan Tetrahydrocannabivarin 9 Tetrahydrocan binol nabinol-C4 Δ -THCV-C3 9 9 nabiorcol Δ -THC-C5 Δ -THC-C4 9 Δ -THCO-C1 9 9 Δ -Tetrahydro- Δ9-Tetrahydro- Δ -Tetrahydro- Δ9-Tetrahydro- cannabinolic -

An Analysis of the Facilitatory Action of Catechol and Phenol at the Neuromuscular Junction of the Cat
Loyola University Chicago Loyola eCommons Dissertations Theses and Dissertations 1972 An Analysis of the Facilitatory Action of Catechol and Phenol at the Neuromuscular Junction of the Cat Joel P. Gallagher Loyola University Chicago Follow this and additional works at: https://ecommons.luc.edu/luc_diss Part of the Medicine and Health Sciences Commons Recommended Citation Gallagher, Joel P., "An Analysis of the Facilitatory Action of Catechol and Phenol at the Neuromuscular Junction of the Cat" (1972). Dissertations. 1138. https://ecommons.luc.edu/luc_diss/1138 This Dissertation is brought to you for free and open access by the Theses and Dissertations at Loyola eCommons. It has been accepted for inclusion in Dissertations by an authorized administrator of Loyola eCommons. For more information, please contact [email protected]. This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 License. Copyright © 1972 Joel P. Gallagher AN ANALYSIS OF THE FACILITATOHY ACTION OF CATECHOL AND PHENOL ·AT THE NEUROMUSCULAR JUNCTION OF THE CAT .A dissertation presented by Joel P. Gallagher for ihe dcg:~ee of DOCTOR OF PHILOSOPHY at the DF.~PARTMENT OF PHARMACOLOGY LOYOLA UNPlERSITY MEDIC.!\L CENTER MAYWOOD, ILLI:;:~OIS F'ebru8.ry, Hl72 lOYOlA ll!RARY UNIViR.SiTY MEDIC "I. "" CENTER ..-------------~----..a"!..... --·-·-·-·--·----~..-----------------------,._,_...,.... ........ __________ -:"I BIOGRAPHY Joel P. Gallagher was born on December 12, 1942, in Chicago, Illinois. In June of 1960 he graduated from Fenwick High School, Oak Park. The following Fall he began his undergraduate training at the College of St. Thomas. St. Paul, Minnesota where he received the Bachelor of II Science degree in Biology-Chemistry in June 1964. -

Cannabinoid Receptor Antagonists Counteract Sensorimotor Gating Deficits in the Phencyclidine Model of Psychosis
Neuropsychopharmacology (2007) 32, 2098–2107 & 2007 Nature Publishing Group All rights reserved 0893-133X/07 $30.00 www.neuropsychopharmacology.org Cannabinoid Receptor Antagonists Counteract Sensorimotor Gating Deficits in the Phencyclidine Model of Psychosis ,1 2,3 4 5 3,6 Martina Ballmaier* , Marco Bortolato , Cristina Rizzetti , Michele Zoli , GianLuigi Gessa , 1,6 4,6 Andreas Heinz and PierFranco Spano 1Department of Psychiatry and Psychotherapy, Charite´ University Medicine, Campus Mitte, Berlin, Germany; 2Department of Pharmacology, 3 4 University of California, Irvine, USA; Department of Neuroscience, Bernard B Brodie, University of Cagliari, Cagliari, Italy; Department of 5 Biomedical Sciences and Biotechnologies, Brescia University Medical School, Brescia, Italy; Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italy Clinical and laboratory findings suggest that cannabinoids and their receptors are implicated in schizophrenia. The role of cannabinoids in schizophrenia remains however poorly understood, as data are often contradictory. The primary aim of this study was to investigate whether the cannabinoid CB1 receptor antagonists rimonabant and AM251 are able to reverse deficits of sensorimotor gating induced by phencyclidine and to mimic the ‘atypical’ antipsychotic profile of clozapine. The prepulse inhibition (PPI) of the startle reflex was used to measure deficits of sensorimotor gating. PPI-disruptive effects of phencyclidine and their antagonism by rimonabant, AM251, and clozapine were studied in rats. The effects of rimonabant were carefully examined taking into account dose ranges, vehicle, and route of administration. We also examined the ability of rimonabant to reduce the PPI-disruptive effects of dizocilpine and apomorphine. Rimonabant as well as AM251 significantly counteracted the phencyclidine-disruptive model of PPI, comparable to the restoring effect of clozapine; no augmentation effect was observed with rimonabant and clozapine as cotreatment.