Population Assessments of Chameleons from Two Montane Sites in Madagascar
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

MADAGASCAR: the Wonders of the “8Th Continent” a Tropical Birding Custom Trip
MADAGASCAR: The Wonders of the “8th Continent” A Tropical Birding Custom Trip October 20—November 6, 2016 Guide: Ken Behrens All photos taken during this trip by Ken Behrens Annotated bird list by Jerry Connolly TOUR SUMMARY Madagascar has long been a core destination for Tropical Birding, and with the opening of a satellite office in the country several years ago, we further solidified our expertise in the “Eighth Continent.” This custom trip followed an itinerary similar to that of our main set-departure tour. Although this trip had a definite bird bias, it was really a general natural history tour. We took our time in observing and photographing whatever we could find, from lemurs to chameleons to bizarre invertebrates. Madagascar is rich in wonderful birds, and we enjoyed these to the fullest. But its mammals, reptiles, amphibians, and insects are just as wondrous and accessible, and a trip that ignored them would be sorely missing out. We also took time to enjoy the cultural riches of Madagascar, the small villages full of smiling children, the zebu carts which seem straight out of the Middle Ages, and the ingeniously engineered rice paddies. If you want to come to Madagascar and see it all… come with Tropical Birding! Madagascar is well known to pose some logistical challenges, especially in the form of the national airline Air Madagascar, but we enjoyed perfectly smooth sailing on this tour. We stayed in the most comfortable hotels available at each stop on the itinerary, including some that have just recently opened, and savored some remarkably good food, which many people rank as the best Madagascar Custom Tour October 20-November 6, 2016 they have ever had on any birding tour. -

Tana Lsms Hh
This PDF generated by katharinakeck, 1/24/2017 10:08:32 AM Sections: 10, Sub-sections: 38, Questionnaire created by opm, 8/4/2016 10:22:56 AM Questions: 366. Last modified by katharinakeck, 1/24/2017 3:00:47 PM Questions with enabling conditions: 206 Questions with validation conditions: 30 Shared with: Rosters: 18 opm (last edited 10/19/2016 10:14:02 AM) Variables: 34 aarau (last edited 10/25/2016 9:18:23 AM) seanoleary (last edited 10/17/2016 4:20:41 PM) arinay (never edited) rharati (never edited) kirsten (never edited) andrianina (never edited) mmihary_r (never edited) sergiy (never edited) janaharb (last edited 10/21/2016 4:55:02 PM) opm (last edited 10/19/2016 10:14:02 AM) gabielte (never edited) TANA_LSMS_HH START Sub-sections: 4, No rosters, Questions: 23, Variables: 5. CONSENT FORM No sub-sections, No rosters, Questions: 1, Static texts: 2. ROSTER No sub-sections, Rosters: 1, Questions: 5, Static texts: 2, Variables: 2. RESPONDENT SELECTION No sub-sections, No rosters, Questions: 7, Variables: 3. MAIN RESPONDENT Sub-sections: 22, Rosters: 10, Questions: 236, Static texts: 4, Variables: 5. CONSUMPTION Sub-sections: 6, Rosters: 5, Questions: 18, Static texts: 4, Variables: 13. HOUSEHOLD HEAD Sub-sections: 2, Rosters: 1, Questions: 18, Static texts: 1, Variables: 3. LABOUR Sub-sections: 4, Rosters: 1, Questions: 42, Variables: 3. OBSERVATIONS No sub-sections, No rosters, Questions: 12. RESULT No sub-sections, No rosters, Questions: 4. APPENDIX A — INSTRUCTIONS APPENDIX B — OPTIONS APPENDIX C — VARIABLES LEGEND 1 / 65 START EA ID NUMERIC: INTEGER ea_id SCOPE: PREFILLED DWELLING ID NUMERIC: INTEGER dwllid SCOPE: PREFILLED TYPE DWELLING ID AGAIN NUMERIC: INTEGER dwllid2 V1 self==dwllid M1 Dwelling ID does not match V2 ea_id*100+1<=self && self <=ea_id*100+30 M2 Dwelling ID and EA ID do not match VARIABLE DOUBLE dwlnum dwllid-100*ea_id THIS IS A REPLACEMENT DWELLING. -
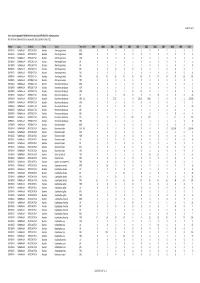
Gross Trade in Appendix II FAUNA (Direct Trade Only), 1999-2010 (For
AC25 Inf. 5 (1) Gross trade in Appendix II FAUNA (direct trade only), 1999‐2010 (for selection process) N.B. Data from 2009 and 2010 are incomplete. Data extracted 1 April 2011 Phylum Class TaxOrder Family Taxon Term Unit 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 Total CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia BOD 0 00001000102 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia BON 0 00080000008 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia HOR 0 00000110406 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia LIV 0 00060000006 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia SKI 1 11311000008 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia SKP 0 00000010001 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia SKU 2 052101000011 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Ammotragus lervia TRO 15 42 49 43 46 46 27 27 14 37 26 372 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Antilope cervicapra TRO 0 00000020002 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae BOD 0 00100001002 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae HOP 0 00200000002 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae HOR 0 0010100120216 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae LIV 0 0 5 14 0 0 0 30 0 0 0 49 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae MEA KIL 0 5 27.22 0 0 272.16 1000 00001304.38 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae MEA 0 00000000101 CHORDATA MAMMALIA ARTIODACTYLA Bovidae Bison bison athabascae -

Final Report
Darwin Initiative – Final Report Darwin project information Project Reference 17-010 Project Title Chameleon trade and conservation in Madagascar Host country(ies) Madagascar UK Contract Holder Durrell Institute of Conservation and Ecology (DICE) UK Partner Institution(s) None Host Country Partner Madagasikara Voakajy Darwin Grant Value £XXX Start/End dates of Project 1 May 2009 to 30 April 2012 Project Leader Name Richard Griffiths Project Website www.madagasikara-voakajy.org https://www.facebook.com/pages/Madagasikara- Voakajy/310507418994236 Report Author(s) and date Richard Jenkins, Christian Randrianantoandro, Richard Griffiths 1 Project Background The island of Madagascar is home to over 83 species of endemic chameleons. These unique lizards, admired by the island’s tourists but disliked by many of its residents, occur in all habitats from sea-level to the highest mountains. However, chameleons in Madagascar are subject to a wide array of threats, including habitat loss, illegal collection and climate change. The three outstanding achievements of this project were: 1) Assessed the conservation status of 76 Malagasy chameleons for the IUCN Red List of Threatened Species 2) Completed a survey of illegal trade in Malagasy chameleons in Thailand 3) Provided essential support to the CITES authorities in Madagascar that led to Standing Committee’s lifting of the trade suspension for some species after 18 years, and an export quota for Furcifer campani 2 Project support to the Convention on Biological Diversity (CBD) (a) This project mainly contributed to two of the Focal Areas in the CBD 2010 Biodiversity Targets. It promoted the conservation of species diversity (Goal 2) by identifying chameleons at highest risk of extinction and then designing and implementing actions to improve the status of these species (Targets 2.1 & 2.2). -

'True Chameleon' from Madagascar: a New, Distinctly Colored
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Zoosystematics and Evolution Jahr/Year: 2018 Band/Volume: 94 Autor(en)/Author(s): Prötzel David, Lambert Shea M., Andrianasolo Ginah Tsiorisoa, Hutter Carl R., Cobb Kerry A., Scherz Mark D., Glaw Frank Artikel/Article: The smallest ‘true chameleon’ from Madagascar: a new, distinctly colored species of the Calumma boettgeri complex (Squamata, Chamaeleonidae) 409-423 Creative Commons Attribution 4.0 licence (CC-BY); original download https://pensoft.net/journals Zoosyst. Evol. 94 (2) 2018, 409–423 | DOI 10.3897/zse.94.27305 The smallest ‘true chameleon’ from Madagascar: a new, distinctly colored species of the Calumma boettgeri complex (Squamata, Chamaeleonidae) David Prötzel1, Shea M. Lambert2, Ginah Tsiorisoa Andrianasolo3, Carl R. Hutter4, Kerry A. Cobb5, Mark D. Scherz1,6, Frank Glaw1 1 Zoologische Staatssammlung München (ZSM-SNSB), Münchhausenstr. 21, 81247 München, Germany 2 Department of Ecology and Evolutionary Biology, University of Arizona, Tucson, AZ 85721, USA 3 Mention Zoologie et Biodiversité Animale, Université d’Antananarivo, BP 906, Antananarivo 101, Madagascar 4 Biodiversity Institute and Department of Ecology and Evolutionary Biology, University of Kansas, Lawrence, KS 66045–7561, USA 5 Department of Biological Sciences, Auburn University, Auburn, AL 36849, USA 6 Zoologisches Institut, Technische Universität Braunschweig, Mendelssohnstr. 4, 38106 Braunschweig, Germany http://zoobank.org/2433A9DD-8AC1-4139-A639-E24053D5C33F Corresponding author: David Prötzel ([email protected]) Abstract Received 8 June 2018 On a recent expedition to eastern Madagascar, we discovered a distinct new species of the Accepted 10 August 2018 genus Calumma that we describe here using an integrative approach combining morphol- Published 19 October 2018 ogy, coloration, osteology and molecular genetics. -

Calumma Vohibola, a New Chameleon Species (Squamata: Chamaeleonidae) from the Littoral Forests of Eastern Madagascar Philip-Sebastian Gehring a , Fanomezana M
This article was downloaded by: [Sebastian Gehring] On: 26 October 2011, At: 23:51 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK African Journal of Herpetology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ther20 Calumma vohibola, a new chameleon species (Squamata: Chamaeleonidae) from the littoral forests of eastern Madagascar Philip-Sebastian Gehring a , Fanomezana M. Ratsoavina a b c , Miguel Vences a & Frank Glaw d a Division of Evolutionary Biology, Zoological Institute, Technical University of Braunschweig, Mendelssohnstr. 4, 38106, Braunschweig, Germany b Département de Biologie Animale, Université d'Antananarivo, BP 906, Antananarivo, 101, Madagascar c Grewcock Center for Conservation Research, Omaha's Henry Doorly Zoo, 3701 South 10th Street, Omaha, NE, 68107-2200, USA d Zoologische Staatssammlung München, Münchhausenstr. 21, 81247, München, Germany Available online: 26 Oct 2011 To cite this article: Philip-Sebastian Gehring, Fanomezana M. Ratsoavina, Miguel Vences & Frank Glaw (2011): Calumma vohibola, a new chameleon species (Squamata: Chamaeleonidae) from the littoral forests of eastern Madagascar, African Journal of Herpetology, 60:2, 130-154 To link to this article: http://dx.doi.org/10.1080/21564574.2011.628412 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and- conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. -

No Longer Single! Description of Female Calumma Vatosoa (Squamata, Chamaeleonidae) Including a Review of the Species and Its Systematic Position
Zoosyst. Evol. 92 (1) 2016, 13–21 | DOI 10.3897/zse.92.6464 museum für naturkunde No longer single! Description of female Calumma vatosoa (Squamata, Chamaeleonidae) including a review of the species and its systematic position David Prötzel1, Bernhard Ruthensteiner1, Frank Glaw1 1 Zoologische Staatssammlung München (ZSM-SNSB), Münchhausenstr. 21, 81247 München, Germany http://zoobank.org/CFD64DFB-D085-4D1A-9AA9-1916DB6B4043 Corresponding author: David Prötzel ([email protected]) Abstract Received 3 September 2015 Calumma vatosoa is a Malagasy chameleon species that has until now been known only Accepted 26 November 2015 from the male holotype and a photograph of an additional male specimen. In this paper Published 8 January 2016 we describe females of the chameleon Calumma vatosoa for the first time, as well as the skull osteology of this species. The analysed females were collected many years before Academic editor: the description of C. vatosoa, and were originally described as female C. linotum. Ac- Johannes Penner cording to external morphology, osteology, and distribution these specimens are assigned to C. vatosoa. Furthermore we discuss the species group assignment of C. vatosoa and transfer it from the C. furcifer group to the C. nasutum group. A comparison of the exter- Key Words nal morphology of species of both groups revealed that C. vatosoa has a relatively shorter distance from the anterior margin of the orbit to the snout tip, more heterogeneous scala- Madagascar tion at the lower arm, a significantly lower number of supralabial and infralabial scales, chameleon and a relatively longer tail than the members of the C. furcifer group. -

CAMELEON Calumma Nasutum (DUMERIL & BIBRON, 1836) DE LA FORET DE MOYENNE ET DE BASSE ALTITUDE DE L’EST DE MADAGASCAR
UNIVERSITE D’ANTANANARIVO FACULTE DES SCIENCES DEPARTEMENT DE BIOLOGIE ANIMALE Latimeria chalumnae MEMOIRE POUR L’OBTENTION DU Diplôme d’Etudes Approfondies (D.E.A.) Formation Doctorale : Sciences de la Vie Option : Biologie, Ecologie et Conservation Animales ESSAI DE CLARIFICATION DE LA TAXONOMIE DU PETIT CAMELEON Calumma nasutum (DUMERIL & BIBRON, 1836) DE LA FORET DE MOYENNE ET DE BASSE ALTITUDE DE L’EST DE MADAGASCAR Présenté par : Mademoiselle Marie Paule RAZAFIMAHATRATRA Devant les membres du Jury composé de : Président : Madame Noromalala RASOAMAMPIONONA RAMINOSOA Professeur d’ESR Rapporteur : Monsieur Daniel RAKOTONDRAVONY Maître de Conférences Examinateurs : Monsieur Achille P. RASELIMANANA Professeur d’ESR Monsieur Hery A. RAKOTONDRAVONY Maître de Conférences Soutenu publiquement le 30 avril 2015 UNIVERSITE D’ANTANANARIVO FACULTE DES SCIENCES DEPARTEMENT DE BIOLOGIE ANIMALE Latimeria chalumnae MEMOIRE POUR L’OBTENTION DU Diplôme d’Etudes Approfondies (D.E.A.) Formation Doctorale : Sciences de la Vie Option : Biologie, Ecologie et Conservation Animales ESSAI DE CLARIFICATION DE LA TAXONOMIE DU PETIT CAMELEON Calumma nasutum (DUMERIL & BIBRON, 1836) DE LA FORET DE MOYENNE ET DE BASSE ALTITUDE DE L’EST DE MADAGASCAR Présenté par : Mademoiselle Marie Paule RAZAFIMAHATRATRA Devant les membres du Jury composé de : Président : Madame Noromalala RASOAMAMPIONONA RAMINOSOA Professeur d’ESR Rapporteur : Monsieur Daniel RAKOTONDRAVONY Maître de Conférences Examinateurs : Monsieur Achille P. RASELIMANANA Professeur d’ESR Monsieur Hery A. -

Toerana Azo Antonina Amin'izany
FAMPAHAFANTARANA Vaccinodrome manerana ny Nosy Ho antsika mpiara-miasa rehetra eto amin’ny Minisiteran’ny Toekarena sy ny Fitantanam-bola, izay maniry ny hanao na eto amin’ny faritra Analamanga na any amin’ny Faritra, dia omena antsika eto ireo toerana azo antonina amin’izany. Faritra Analamanga ATSINANANA TOAMASINA I CHUM TOAMASINA CSB Urbain AMBOHIDRATRIMO TOAMASINA II BSD TOAMASINA II CSB 2 Ivato Aeroport, CSB 2 Mahitsy, CSB 2 Antehiroka, CSB 2 CSB FANANDRANA Soavinimerina, CSB 2 Talatamaty, CSB 2 Ambohitrimanjaka, CSB FOULPOINTE CSB 2 Ambodirano, CSB 2 Ampangabe, CSB 2 Ambatolampy BRICKAVILLE CSB URBAIN BRICKAVILLE tsimahafotsy, CSB 2 Antanetibe Mahazaza, CSB 2 Anosiala. CHRD2 BRICKAVILLE MAHANORO CHRD MAHANORO ANTANANARIVO ATSIMONDRANO CSB U MAHANORO Hopitaly Itaosy, CSB 2 Andoharanofotsy, CSB 2 Ankadivoribe, CSB1 AMPITAKIHOSY CSB 2 Faliarivo, CSB 2 Itaosy Bemasoandro, CSB 2 Anosizato, VATOMANDRY BSD VATOMANDRY CSB 2 Itaosy Cité, CSB 2 Anjomakely, CSB 2 Tanjombato, CSB URBAIN VATOMANDRY CSB 2 Fenoarivo, CSB 2 Tsararivotra, CSB 2 Soavina, CSB 2 MAROLAMBO CHRD MAROLAMBO Ambohijafy, CSB 2 Ankaraobato, CSB 2 Ambohijanaka, CSB 2 CSB2 MAROLAMBO Ambaniala Itaosy. ANTANAMBAO MANAMPONTSY CHRD ANTANAMBAO MANAMPOTSY ANTANANARIVO AVARADRANO CSB 2 Anosiavaratra, CSB 2 Ankadikely, CSB 2 Ampahimanga, DIANA CSB 2 Masindray, CSB 2 Ambohimangakely, CSB 2 Alasora, ANTSIRANANA 1 CSBU TANAMBAO CSB 2 Sabotsy Namehana, CSB 2 Talata Volonondry, CSB 2 AMBILOBE CSB2 AMBILOBE Anjeva, CSB 2 Ankadinandrina, CSB 2 Ambohimanga Rova, CSB2 AMPONDRALAVA CSB 2 Ambohimalaza. -

Habitat Use and Abundance of a Low-Altitude Chameleon Assemblage in Eastern Madagascar
HERPETOLOGICAL JOURNAL 17: 247–254, 2007 Habitat use and abundance of a low-altitude chameleon assemblage in eastern Madagascar Jeanneney Rabearivony1, Lee D. Brady2, Richard K.B. Jenkins3,4 & Olga R. Ravoahangimalala1 1Département de Biologie Animale, Université d’Antananarivo, Madagascar 2Durrell Institute of Conservation and Ecology, University of Kent, Canterbury, UK 3School of Biological Sciences, University of Aberdeen, UK 4Madagasikara Voakajy, Antananarivo, Madagascar We studied the density and abundance of chameleons in a lowland Malagasy rainforest during the austral summer and winter. Nocturnal searches for chameleons were conducted along transects within relatively intact forest and vegetation on abandoned agricultural land adjacent to the forest. Four chameleon species were encountered during the study, Brookesia superciliaris, Calumma parsonii parsonii, Calumma nasutum and Furcifer pardalis. Brookesia superciliaris was most common inside relatively intact forest and the few individuals located in the regenerating forest on abandoned agricultural land were found in tiny, isolated patches of degraded rainforest next to rivers. Calumma p. parsonii was only encountered on three occasions in relatively intact forest and was a rare member of the community. The abundance of C. nasutum was highest in relatively intact forest but this species also occurred in vegetation on abandoned agricultural land. Furcifer pardalis was only found on the abandoned agricultural land, where it was observed laying eggs in sandy soil in August. The abundance of all species in habitats alongside rivers was higher in January than July–August, with the exception of C. p. parsonii, which was not detected during the former period. Additional investigations into habitat preference of chameleons and surveys in other forests in region are needed to establish whether the low abundance of C. -

A Phylogeny and Revised Classification of Squamata, Including 4161 Species of Lizards and Snakes
BMC Evolutionary Biology This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted PDF and full text (HTML) versions will be made available soon. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes BMC Evolutionary Biology 2013, 13:93 doi:10.1186/1471-2148-13-93 Robert Alexander Pyron ([email protected]) Frank T Burbrink ([email protected]) John J Wiens ([email protected]) ISSN 1471-2148 Article type Research article Submission date 30 January 2013 Acceptance date 19 March 2013 Publication date 29 April 2013 Article URL http://www.biomedcentral.com/1471-2148/13/93 Like all articles in BMC journals, this peer-reviewed article can be downloaded, printed and distributed freely for any purposes (see copyright notice below). Articles in BMC journals are listed in PubMed and archived at PubMed Central. For information about publishing your research in BMC journals or any BioMed Central journal, go to http://www.biomedcentral.com/info/authors/ © 2013 Pyron et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes Robert Alexander Pyron 1* * Corresponding author Email: [email protected] Frank T Burbrink 2,3 Email: [email protected] John J Wiens 4 Email: [email protected] 1 Department of Biological Sciences, The George Washington University, 2023 G St. -

The Project on Master Plan Formulation for Economic Axis of Tatom (Antananarivo-Toamasina, Madagasikara)
Ministry of Regional Development, Building, Housing and Public Works (MAHTP) Government of the Republic of Madagascar The Project on Master Plan Formulation for Economic Axis of TaToM (Antananarivo-Toamasina, Madagasikara) Final Report Summary October 2019 Japan International Cooperation Agency (JICA) Oriental Consultants Global Co., Ltd. CTI Engineering International Co., Ltd. CTI Engineering Co., Ltd. EI JR 19-102 Ministry of Regional Development, Building, Housing and Public Works (MAHTP) Government of the Republic of Madagascar The Project on Master Plan Formulation for Economic Axis of TaToM (Antananarivo-Toamasina, Madagasikara) Final Report Summary October 2019 Japan International Cooperation Agency (JICA) Oriental Consultants Global Co., Ltd. CTI Engineering International Co., Ltd. CTI Engineering Co., Ltd. Currency Exchange Rates EUR 1.00 = JPY 127.145 EUR 1.00 = MGA 3,989.95 USD 1.00 = JPY 111.126 USD 1.00 = MGA 3,489.153 MGA 1.00 = JPY 0.0319 Average during the period between June 2018 and June 2019 Administrative Divisions of Madagascar The decentralised administrative divisions of Madagascar is divided into 22 regions which are further divided into 114 districts. The districts are further divided into communes and each communes into fokontany. Besides the decentralised administrative divisions, the country is subdivided into six provinces, divided into 24 prefectures. The prefectures are divided into 117 districts and further into arrondissements. The boundary of region and prefecture are same except for two prefectures Nosy