The Earliest Fossil Record of the Poorly Known Family Condylocardiidae from Argentina
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pineda-Salgado, 2020 CD
Universidad Nacional de La Plata Facultad de Ciencias Naturales y Museo Análisis de concentraciones fósiles en la Formación Monte León (Mioceno inferior), en la costa de la provincia de Santa Cruz Gabriela Pineda Salgado Tesis doctoral Directores: Miguel Griffin, Ana Parras 2020 A mi mamá, mi abuela y mi abuelichi A Señor Pantufla y Spock-Uhura, los félidos con más estilo Agradecimientos Al Posgrado de la Facultad de Ciencias Naturales y Museo de la Universidad Nacional de La Plata. A los doctores Miguel Griffin y Ana Parras por dirigirme, por todas las facilidades brindadas para la realización de este trabajo, así como por su ayuda en las labores de campo y por los apoyos obtenidos para exponer parte de los resultados del mismo en reuniones nacionales e internacionales. Al jurado conformado por los doctores Claudia del Río, Miguel Manceñido y Sven Nielsen. A la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) por la beca doctoral otorgada a través del Fondo para la Investigación Científica y Tecnológica (FONCyT), en el marco del Proyecto de Investigación Científica y Tecnológica PICT 2012-1726. Al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) por la beca interna de finalización de doctorado otorgada durante el periodo 2017-2019. Al Instituto de Ciencias de la Tierra y Ambientales de La Pampa (INCITAP, CONICET- UNLPam) y a la Facultad de Ciencias Exactas y Naturales de la UNLPam por ceder el espacio institucional para el desarrollo de esta tesis. A la Administración de Parques Nacionales por autorizar la recolección de muestras en los límites del Parque Nacional Monte León. -

Marine Bivalve Molluscs
Marine Bivalve Molluscs Marine Bivalve Molluscs Second Edition Elizabeth Gosling This edition first published 2015 © 2015 by John Wiley & Sons, Ltd First edition published 2003 © Fishing News Books, a division of Blackwell Publishing Registered Office John Wiley & Sons, Ltd, The Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, UK Editorial Offices 9600 Garsington Road, Oxford, OX4 2DQ, UK The Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, UK 111 River Street, Hoboken, NJ 07030‐5774, USA For details of our global editorial offices, for customer services and for information about how to apply for permission to reuse the copyright material in this book please see our website at www.wiley.com/wiley‐blackwell. The right of the author to be identified as the author of this work has been asserted in accordance with the UK Copyright, Designs and Patents Act 1988. All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, except as permitted by the UK Copyright, Designs and Patents Act 1988, without the prior permission of the publisher. Designations used by companies to distinguish their products are often claimed as trademarks. All brand names and product names used in this book are trade names, service marks, trademarks or registered trademarks of their respective owners. The publisher is not associated with any product or vendor mentioned in this book. Limit of Liability/Disclaimer of Warranty: While the publisher and author(s) have used their best efforts in preparing this book, they make no representations or warranties with respect to the accuracy or completeness of the contents of this book and specifically disclaim any implied warranties of merchantability or fitness for a particular purpose. -

Paleontology and Sedimentology of Middle Eocene Rocks in Lago Argentino Area, Santa Cruz Province, Argentina
AMEGHINIANA (Rev. Asoc. Paleontol. Argent.) - 46 (1): 27-47. Buenos Aires, 30-04-2009 ISSN 0002-7014 Paleontology and sedimentology of Middle Eocene rocks in Lago Argentino area, Santa Cruz Province, Argentina Silvio CASADÍO1,2, Miguel GRIFFIN1,2, Sergio MARENSSI3,2, Laura NET4, Ana PARRAS1,2, Martín RODRÍGUEZ RAISING5,2 and Sergio SANTILLANA3 Abstract. Sedimentological and paleontological study of the Man Aike Formation at the Estancia 25 de Mayo, SW of Santa Cruz Province, Argentina, represents the evolution of an incised valley from fluvial to marine environment during the late middle Eocene. At the base of the unit there is an unconformity that corresponds to fluvial channels which cut down into the underlying Maastrichtian sandstones of the Calafate Formation. The fauna of invertebrates (mostly molluscs) illustrated herein was collected from shell beds interpreted as tidal ravinement surfaces. The fauna includes terebratulid brachiopods, bivalves of the families Malletiidae, Mytilidae, Pinnidae, Ostreidae, Pectinidae, Carditidae, Crassatellidae, Lahillidae, Mactridae, Veneridae, and Hiatellidae, and gastropods of the families Trochidae and Calyptraeidae, and a member of Archaeogastropoda of uncertain affinities. The similarities of this fauna with that recorded in the Upper Member of the Río Turbio Formation, together with 87Sr/86Sr ages, sug- gest a late Middle Eocene age for the Man Aike Formation. Resumen. PALEONTOLOGÍA Y SEDIMENTOLOGÍA DE LAS ROCAS DEL EOCENO MEDIO EXPUESTAS EN EL ÁREA DE LAGO ARGENTINO, PROVINCIA DE SANTA CRUZ, ARGENTINA. Los estudios sedimentológicos y paleontológicos real- izados en rocas asignadas a la Formación Man Aike, expuestas en la estancia 25 de Mayo, al sur de Calafate, provincia de Santa Cruz, Argentina, sugieren que esta unidad representa la evolución de un valle inciso desde ambientes fluviales a marinos durante el Eoceno medio tardío. -
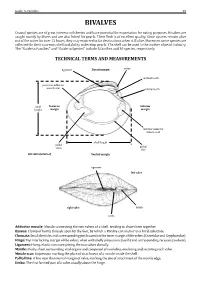
Field Identification Guide to the Living Marine Resources In
Guide to Families 29 BIVALVES Coastal species are of great interest to fisheries and have potential for exportation for eating purposes. Bivalves are caught mainly by divers and are also fished for pearls. Their flesh is of excellent quality. Since oysters remain alive out of the water for over 12 hours, they may exported to far destinations when still alive. Moreover, some species are collected for their nacreous shell and ability to develop pearls. The shell can be used in the mother of pearl industry. The “Guide to Families’’ andTECHNICAL ‘‘Guide to Species’’ TERMS include 5AND families MEASUREMENTS and 10 species, respectively. ligament Dorsal margin umbo posterior adductor cardinal tooth muscle scar lateral tooth Posterior Anterior margin margin shell height anterior adductor muscle scar pallial sinus pallial shell length line left valve (interior) Ventral margin ligament left valve right valve lunule umbo Adductor muscle: Byssus: Chomata: Muscle connecting the two valves of a shell, tending to draw them together. Hinge: Clump of horny threads spun by the foot, by which a Bivalve can anchor to a hard substrate. Ligament: Small denticles and corresponding pits located on the inner margin of the valves (Ostreidae and Gryphaeidae). Mantle: Top interlocking margin of the valves, often with shelly projections (teeth) and corresponding recesses (sockets). Muscle scar: Horny, elastic structure joining the two valves dorsally. Pallial line: Fleshy sheet surrounding vital organs and composed of two lobes, one lining and secreting each valve. Umbo: Impression marking the place of attachment of a muscle inside the shell. A line near the internal margin of valve, marking the site of attachment of the mantle edge. -

The Recent Molluscan Marine Fauna of the Islas Galápagos
THE FESTIVUS ISSN 0738-9388 A publication of the San Diego Shell Club Volume XXIX December 4, 1997 Supplement The Recent Molluscan Marine Fauna of the Islas Galapagos Kirstie L. Kaiser Vol. XXIX: Supplement THE FESTIVUS Page i THE RECENT MOLLUSCAN MARINE FAUNA OF THE ISLAS GALApAGOS KIRSTIE L. KAISER Museum Associate, Los Angeles County Museum of Natural History, Los Angeles, California 90007, USA 4 December 1997 SiL jo Cover: Adapted from a painting by John Chancellor - H.M.S. Beagle in the Galapagos. “This reproduction is gifi from a Fine Art Limited Edition published by Alexander Gallery Publications Limited, Bristol, England.” Anon, QU Lf a - ‘S” / ^ ^ 1 Vol. XXIX Supplement THE FESTIVUS Page iii TABLE OF CONTENTS INTRODUCTION 1 MATERIALS AND METHODS 1 DISCUSSION 2 RESULTS 2 Table 1: Deep-Water Species 3 Table 2: Additions to the verified species list of Finet (1994b) 4 Table 3: Species listed as endemic by Finet (1994b) which are no longer restricted to the Galapagos .... 6 Table 4: Summary of annotated checklist of Galapagan mollusks 6 ACKNOWLEDGMENTS 6 LITERATURE CITED 7 APPENDIX 1: ANNOTATED CHECKLIST OF GALAPAGAN MOLLUSKS 17 APPENDIX 2: REJECTED SPECIES 47 INDEX TO TAXA 57 Vol. XXIX: Supplement THE FESTIVUS Page 1 THE RECENT MOLLUSCAN MARINE EAUNA OE THE ISLAS GALAPAGOS KIRSTIE L. KAISER' Museum Associate, Los Angeles County Museum of Natural History, Los Angeles, California 90007, USA Introduction marine mollusks (Appendix 2). The first list includes The marine mollusks of the Galapagos are of additional earlier citations, recent reported citings, interest to those who study eastern Pacific mollusks, taxonomic changes and confirmations of 31 species particularly because the Archipelago is far enough from previously listed as doubtful. -

FAO Fisheries & Aquaculture
Food and Agriculture Organization of the United Nations Fisheries and for a world without hunger Aquaculture Department Cultured Aquatic Species Information Programme Mercenaria mercenaria (Linnaeus, 1758) I. Identity V. Status And Trends a. Biological Features VI. Main Issues b. Images Gallery a. Responsible Aquaculture Practices II. Profile VII. References a. Historical Background a. Related Links b. Main Producer Countries c. Habitat And Biology III. Production a. Production Cycle b. Production Systems c. Diseases And Control Measures IV. Statistics a. Production Statistics b. Market And Trade Identity Mercenaria mercenaria Linnaeus, 1758 [Veneridae] FAO Names: En - Northern quahog(=Hard clam), Fr - Praire, Es - Chirla mercenaria Biological features Shell solid, equivalve; inequilateral, beaks in the front half of the shell; broadly oval in outline. Ligament a deeply inset, dark brown elliptical band, behind the beaks reaching half-way to the posterior margin. Lunule well defined, broad, heart-shaped. Escutcheon indistinct. Sculpture of concentric lines, raised here and there into ridges, and fine radiating lines. In young specimens the ridges are present all over the shell but in the adult they persist, after wear and tear, only near the anterior and posterior margins. Growth stages prominent. Both valves with three cardinal teeth; in addition there is present in each valve a rough tooth-like area behind the beaks and immediately below the ligament; this area has the appearance of a supplementary posterior cardinal tooth which has been broken off. No laterals. Pallial sinus not deep, triangular. Margin grenulate. Colour a dirty white, light varnish-brown, dull grey or grey-brown. Inside of shell white, sometimes deep violet about the adductor muscle scars. -

List of Bivalve Molluscs from British Columbia, Canada
List of Bivalve Molluscs from British Columbia, Canada Compiled by Robert G. Forsyth Research Associate, Invertebrate Zoology, Royal BC Museum, 675 Belleville Street, Victoria, BC V8W 9W2; [email protected] Rick M. Harbo Research Associate, Invertebrate Zoology, Royal BC Museum, 675 Belleville Street, Victoria BC V8W 9W2; [email protected] Last revised: 11 October 2013 INTRODUCTION Classification rankings are constantly under debate and review. The higher classification utilized here follows Bieler et al. (2010). Another useful resource is the online World Register of Marine Species (WoRMS; Gofas 2013) where the traditional ranking of Pteriomorphia, Palaeoheterodonta and Heterodonta as subclasses is used. This list includes 237 bivalve species from marine and freshwater habitats of British Columbia, Canada. Marine species (206) are mostly derived from Coan et al. (2000) and Carlton (2007). Freshwater species (31) are from Clarke (1981). Common names of marine bivalves are from Coan et al. (2000), who adopted most names from Turgeon et al. (1998); common names of freshwater species are from Turgeon et al. (1998). Changes to names or additions to the fauna since these two publications are marked with footnotes. Marine groups are in black type, freshwater taxa are in blue. Introduced (non-indigenous) species are marked with an asterisk (*). Marine intertidal species (n=84) are noted with a dagger (†). Quayle (1960) published a BC Provincial Museum handbook, The Intertidal Bivalves of British Columbia. Harbo (1997; 2011) provided illustrations and descriptions of many of the bivalves found in British Columbia, including an identification guide for bivalve siphons and “shows”. Lamb & Hanby (2005) also illustrated many species. -

Proceedings of the Academy of Natural Sciences of Philadelphia
32 PROCEEDINGS OF THE ACADEMY OF [1887. ON NEW GENEEIC FOKMS OF CRETACEOUS MOLLUSCA AND THEIE RELATION TO OTHER FORMS. BY CHARLES A. WHITE. Published by permission of the Director of the United States Geological Survey. The type species of the three generic forms which are described in this article ^ belong to the collections of Cretaceous fossils from Texas, which I am now preparing for publication in one of the memoirs of the U. S. Geological Survey. In their generic charac- teristics all three of them appear to be respectively identical with certain forms which have long been known, but which have been referred to other genera by different authors. The features which I now present as having generic value seem to have been overlooked by those authors, or, so far as they were observed, they were treated as specific characters. Two of these forms belong to the section Melininse of the family Aviculidse. The other is referred to the Crassatellidse, but it departs considerably from the typical section of that family. CRASSATELLIDJE. Genus STEARNSIA (gen. nov.). Shell compressed, subtrihedral or subcircular in marginal out- line; beaks small, closely approximate, prominent by reason of the abrupt sloping away of both the antero-and postero-dorsal borders; lunule and escutcheon both well defined and flattened or excavated; hinge strong, consisting of both cardinal and lateral teeth; cardinal teeth two in the left valve and three in the right; both posterior and anterior lateral teeth long and slender; posterior laterals two in the right valve and one in the left; anterior laterals two in the left valve and one in the right. -

Spissatella Trailli (Lectotype TM2859 – Awamoa, Oamaru, New Zealand)
JOURNEYS THROUGH SHAPE AND TIME: PALAEOBIOLOGY OF CENOZOIC NEW ZEALAND SPISSATELLA AND EUCRASSATELLA (BIVALVIA, CRASSATELLIDAE) by Katie Susanna Collins A thesis submitted to the Victoria University of Wellington in fulfillment of the requirements for the degree of Doctor of Philosophy Victoria University of Wellington 2013 Spissatella trailli (Lectotype TM2859 – Awamoa, Oamaru, New Zealand) i ii ABSTRACT A novel, highly-integrated approach combining morphometric, stratocladistic and sclerochronological methods has been applied to two genera of New Zealand Cenozoic crassatellid bivalve (Family Crassatellidae): Spissatella Finlay, 1926 and Eucrassatella Iredale, 1924. This study builds on previous work on Spissatella that demonstrated their amenability to shape analysis and provided a foundation for evolutionary studies of the group. The taxonomy of these crassatellids has been in need of revision; a number of changes to generic placement having been proposed in recent publications without redescription. These bivalves are character-depauperate and known only from fossil material within New Zealand, making them challenging subjects for the phylogenetic analysis that would, ideally, inform taxonomic revision. Geometric morphometric methods have been used to characterise the morphological variation of the study group in terms of shape. Landmarks/semilandmarks that capture internal hard-part morphology and external shell shape, have been compared with internal landmarks only, outline shape semilandmarks only, and outline shape Fourier transform methods, and are shown to best combine comprehensive coverage of total shell form with high correct reassignment of individuals to taxa in multidimensional morphospace. Procrustes- superimposed landmark/semilandmark configurations have been ordinated using Principal Components Analysis (PCA), and PCA plots have been used to compare the shape variation of each species. -

Three Alien Molluscs from Iskenderun Bay (SE Turkey)
Aquatic Invasions (2006) Volume 1, Issue 2: 76-79 DOI 10.3391/ai.2006.1.2.4 © 2006 The Author(s) Journal compilation © 2006 REABIC (http://www.reabic.net) This is an Open Access article Research article Three alien molluscs from Iskenderun Bay (SE Turkey) Doğan Çeviker1 and Serhat Albayrak2* 1Itri Sokak No:2 34349 Balmumcu-Istanbul, Turkey E-mail: [email protected] 2Istanbul University, Faculty of Science, Department of Biology 34118 Vezneciler-Istanbul, Turkey E-mail: [email protected] *Corresponding author Received 26 April 2006; accepted in revised form 4 May 2006 Abstract This study reports the presence of three alien molluscs from Iskenderun Bay (SE Turkey). Amathina tricarinata (Linnaeus, 1767) and Petricola hemprichi Issel, 1869 have prior records from other regions of Mediterranean, but, Cardites akabana (Sturany, 1899) first recorded in this paper. Since all of them are present in the Red Sea or Suez Canal, they can be considered as Lessepsian immigrants. Key words: Mollusca, alien species, Mediterranean, Turkey Introduction that 88 % of the exotic molluscs are Lessepsian immigrants in the eastern Mediterranean (Galil The Mediterranean Sea hosts about 8500 species and Zenetos 2002). Detailed data about these species of macroscopic animals. This rich biodiversity, are available on the Internet (www.ciesm.org/atlas). representing 8-9 % of total species number of the Either Lessepsian or non-Lessepsian, many world’s seas, comprises temperate and sub- new non-indigenous species continue to enter the tropical elements together with endemic and Mediterranean. alien species (Zenetos et al. 2002). The eastern Mediterranean is most vulnerable The introduction of alien species (also known to invasion and should be continuously as exotic, introduced or non-native species) into monitored. -

TREATISE ONLINE Number 48
TREATISE ONLINE Number 48 Part N, Revised, Volume 1, Chapter 31: Illustrated Glossary of the Bivalvia Joseph G. Carter, Peter J. Harries, Nikolaus Malchus, André F. Sartori, Laurie C. Anderson, Rüdiger Bieler, Arthur E. Bogan, Eugene V. Coan, John C. W. Cope, Simon M. Cragg, José R. García-March, Jørgen Hylleberg, Patricia Kelley, Karl Kleemann, Jiří Kříž, Christopher McRoberts, Paula M. Mikkelsen, John Pojeta, Jr., Peter W. Skelton, Ilya Tëmkin, Thomas Yancey, and Alexandra Zieritz 2012 Lawrence, Kansas, USA ISSN 2153-4012 (online) paleo.ku.edu/treatiseonline PART N, REVISED, VOLUME 1, CHAPTER 31: ILLUSTRATED GLOSSARY OF THE BIVALVIA JOSEPH G. CARTER,1 PETER J. HARRIES,2 NIKOLAUS MALCHUS,3 ANDRÉ F. SARTORI,4 LAURIE C. ANDERSON,5 RÜDIGER BIELER,6 ARTHUR E. BOGAN,7 EUGENE V. COAN,8 JOHN C. W. COPE,9 SIMON M. CRAgg,10 JOSÉ R. GARCÍA-MARCH,11 JØRGEN HYLLEBERG,12 PATRICIA KELLEY,13 KARL KLEEMAnn,14 JIřÍ KřÍž,15 CHRISTOPHER MCROBERTS,16 PAULA M. MIKKELSEN,17 JOHN POJETA, JR.,18 PETER W. SKELTON,19 ILYA TËMKIN,20 THOMAS YAncEY,21 and ALEXANDRA ZIERITZ22 [1University of North Carolina, Chapel Hill, USA, [email protected]; 2University of South Florida, Tampa, USA, [email protected], [email protected]; 3Institut Català de Paleontologia (ICP), Catalunya, Spain, [email protected], [email protected]; 4Field Museum of Natural History, Chicago, USA, [email protected]; 5South Dakota School of Mines and Technology, Rapid City, [email protected]; 6Field Museum of Natural History, Chicago, USA, [email protected]; 7North -

Mollusca, Bivalvia): Protocardiinae, Laevicardiinae, Lahilliinae, Tulongocardiinae Subfam
Zoologica Scripta, Vol. 24, No. 4, pp. 321-346, 1995 Pergamon Elsevier Science Ltd The Norwegian Academy of Science and Letters Printed in Great Britain 0300-3256(95)00011-9 Phytogeny of the Cardiidae (Mollusca, Bivalvia): Protocardiinae, Laevicardiinae, Lahilliinae, Tulongocardiinae subfam. n. and Pleuriocardiinae subfam. n. JAY A. SCHNEIDER Accepted 7 June 1995 Schneider, J.A. 1995. Phytogeny of the Cardiidae (Mollusca, Bivalvia): Protocardiinae, Laevicar- diinae, Lahilliinae, Tulongocardiinae subfam.n. and Pleuriocardiinae subfam.n.—Zool. Scr. 24: 321-346. In a preliminary cladistic analysis of the bivalve family Cardiidae (Schneider 1992), members of the subfamilies Protocardiinae, Lahilliinae, and Laevicardiinae, plus the genus Nemocardium, were found to be the least derived taxa of cardiids. A cladistic analysis is undertaken of the genera and subgenera of these cardiid taxa, plus several Mesozoic taxa which have never been assigned to any subfamily. The Late Triassic Tulongocardium, which is placed in Tulongocardiinae subfam. n., is the sister taxon to all other cardiids. Protocardiinae is restricted to the genus Protocardia. Most other Mesozoic taxa which have been placed in the Protocardiinae are found to be members of the Lahilliinae. Nemocardium is placed in the Laevicardiinae. Incacardium, Pleuriocardia, and Dochmocardia form a monophyletic group, Pleuriocardiinae subfam. n. Pleuriocardiinae, Laevi- cardiinae, and the remaining members of the Cardiidae (herein informally termed "cucardiids") form a monophyletic group. Jay A. Schneider, Smithsonian Tropical Research Institute, Box2072, Balboa, Republic of Panama. Present address: Department of Geology, Youngstown State University, Youngstown, OH 44555- 3672, C/.S./l. re7..2;6-742-77JJ;Fa%.276-742-/7J4 Introduction Bivalves of the family Cardiidae (cockles and giant clams) Outgroup Palaeocardita originated in the Late Triassic and have a present-day Septocardia diversity of nearly 200 species (Rosewater 1965; Fischer- Protocardia | PROTOCARDIINAE Integricardium I Piette 1977).