Eoursivivas Cultriformis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Greening Phenomenon in Bivalve by Marennine Produced from Haslea Ostrearia and Its Consequences on Bivalve's Integrated Resp
Greening phenomenon in bivalve by marennine produced from Haslea ostrearia and its consequences on bivalve’s integrated response Fiddy Semba Prasetiya To cite this version: Fiddy Semba Prasetiya. Greening phenomenon in bivalve by marennine produced from Haslea os- trearia and its consequences on bivalve’s integrated response. Invertebrate Zoology. Université du Maine, 2015. English. NNT : 2015LEMA1017. tel-01279527 HAL Id: tel-01279527 https://tel.archives-ouvertes.fr/tel-01279527 Submitted on 26 Feb 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Fiddy SEMBA PRASETIYA Mémoire présenté en vue de l’obtention du grade de Docteur de l’Université du Maine sous le label de L’Université Nantes Angers Le Mans École doctorale : Végétale Environnement Nutrition Agro-alimentaire Mer (VENAM) Discipline : BIOLOGIÉ DES ORGANISMES Unité de recherche : MER MOLÉCULE ET SANTÉ (MMS) – EA n°2160, Université du Maine, UFR Sciences et Techniques, Avenue Olivier Messiaen 72085 Le Mans Cedex 9 Soutenue le 27 Novembre 2015 Greening phenomenon in bivalve by marennine -

Larvae of Bivalve Mollusks of the Sevastopol Region of the Black Sea
W&M ScholarWorks Reports 1966 Larvae of bivalve mollusks of the Sevastopol region of the Black Sea K. A. Zakhvatkina Follow this and additional works at: https://scholarworks.wm.edu/reports Part of the Aquaculture and Fisheries Commons, Marine Biology Commons, and the Zoology Commons Recommended Citation Zakhvatkina, K. A. (1966) Larvae of bivalve mollusks of the Sevastopol region of the Black Sea. Translation series (Virginia Institute of Marine Science) ; no. 15. Virginia Institute of Marine Science, William & Mary. https://scholarworks.wm.edu/reports/39 This Report is brought to you for free and open access by W&M ScholarWorks. It has been accepted for inclusion in Reports by an authorized administrator of W&M ScholarWorks. For more information, please contact [email protected]. VIRGINIA INSTITUTE OF MARINE SCIENCE GLOUCESTER POINT, VIRGDHA .. LARVAE OF BIVALVE MOLLUSKS OF THE SEVASTOPOL REGION OF THE BLACK SEA TRANSlATION SLRIES NO· 15 1966 Virginia Institute of Marine Science Gloucester Point, Virginia URVhE OF BIVALVE hOLLUSKS OF THE SEVii.STOPOL REGION OF THE BLACK SEA By K. A· Zakhvatkina Original title: Lichinki dvustvorchatykh molliuskov sevastopol'skogo raiona Chernogo Moria From: Akademiia Nauk SSSR, Trudy Sevastopol1 skoi Biologicheskoi Stantsii, Tom XI, p• 108-152, 1959 Translated by Evelyn c. wells Edited by Paul Chanley TRANSLATION SERIES NO· 15 w. J. Hargis Director April 1966 Akad • Nauk SSSR, Trudy Sevastopol' skoi Biologicheskoi St.antsii Tom XI, P• 108-152, 1959 LJ.dtvii.E OF BIV;.,LVE HOLLUSKS. OF TH1 SLVJL.TOf'OL REGION OF THE BLACK SEA By K· A· Zakhvatkina Heretofore, the systematic relationships as well as the biology p•l08 and ecology of bivalve larvce have been poorly known• The work of A· Borisiak (1905), on the larv~e of bivalve mollusks, is of only historical interest since only four of the 20 forfus described were deter~mined to genus• Data on the reproduction of several species of bivalve mollusks, especially on spawning seasons, are given in the work of z. -
Field Identification Guide to the Living Marine Resources In
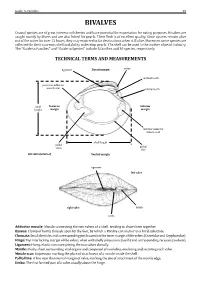
Guide to Families 29 BIVALVES Coastal species are of great interest to fisheries and have potential for exportation for eating purposes. Bivalves are caught mainly by divers and are also fished for pearls. Their flesh is of excellent quality. Since oysters remain alive out of the water for over 12 hours, they may exported to far destinations when still alive. Moreover, some species are collected for their nacreous shell and ability to develop pearls. The shell can be used in the mother of pearl industry. The “Guide to Families’’ andTECHNICAL ‘‘Guide to Species’’ TERMS include 5AND families MEASUREMENTS and 10 species, respectively. ligament Dorsal margin umbo posterior adductor cardinal tooth muscle scar lateral tooth Posterior Anterior margin margin shell height anterior adductor muscle scar pallial sinus pallial shell length line left valve (interior) Ventral margin ligament left valve right valve lunule umbo Adductor muscle: Byssus: Chomata: Muscle connecting the two valves of a shell, tending to draw them together. Hinge: Clump of horny threads spun by the foot, by which a Bivalve can anchor to a hard substrate. Ligament: Small denticles and corresponding pits located on the inner margin of the valves (Ostreidae and Gryphaeidae). Mantle: Top interlocking margin of the valves, often with shelly projections (teeth) and corresponding recesses (sockets). Muscle scar: Horny, elastic structure joining the two valves dorsally. Pallial line: Fleshy sheet surrounding vital organs and composed of two lobes, one lining and secreting each valve. Umbo: Impression marking the place of attachment of a muscle inside the shell. A line near the internal margin of valve, marking the site of attachment of the mantle edge. -

Clam Dissection Guideline
Clam Dissection Guideline BACKGROUND: Clams are bivalves, meaning that they have shells consisting of two halves, or valves. The valves are joined at the top, and the adductor muscles on each side hold the shell closed. If the adductor muscles are relaxed, the shell is pulled open by ligaments located on each side of the umbo. The clam's foot is used to dig down into the sand, and a pair of long incurrent and excurrent siphons that extrude from the clam's mantle out the side of the shell reach up to the water above (only the exit points for the siphons are shown). Clams are filter feeders. Water and food particles are drawn in through one siphon to the gills where tiny, hair-like cilia move the water, and the food is caught in mucus on the gills. From there, the food-mucus mixture is transported along a groove to the palps (mouth flaps) which push it into the clam's mouth. The second siphon carries away the water. The gills also draw oxygen from the water flow. The mantle, a thin membrane surrounding the body of the clam, secretes the shell. The oldest part of the clam shell is the umbo, and it is from the hinge area that the clam extends as it grows. I. Purpose: The purpose of this lab is to identify the internal and external structures of a mollusk by dissecting a clam. II. Materials: 2 pairs of safety goggles 1 paper towel 2 pairs of gloves 1 pair of scissors 1 preserved clam 2 pairs of forceps 1 dissecting tray 2 probes III. -

What Are Molluscs?
The shells of molluscs from all over the world – on land, in lakes, and in the ocean – contain very detailed imprints of past climate change. Using isotope analysis, we can extract these signals and start to piece together long-term climate variations. You will never look at a garden snail in the same way again! What are molluscs? Molluscs are soft-bodied (invertebrate) organisms that are widespread in terrestrial, freshwater, and marine habitats. We can split them into two basic groups: Gastropods: Molluscs with up to one shell or ‘valve’ (such as snails or slugs) Bivalves: Molluscs with two-sided shells or ‘valves’ (such as clams, oysters, and mussels) Many molluscs (both gastropods and bivalves) build hard shells that are rich in calcium carbonate. We call these ‘calcareous shells’. Such shells are formed in distinct bands (like tree rings) growing outwards along the direction of growth – the oldest shell is at the edge. Over time, more bands are added to the shell as it grows and the organism is enlarged (Figure 1a-c). The environmental conditions in which the organism live are reflected in the types of isotopes that are present in its shell. This means that the shell contains a ‘geochemical’ signature of the mollusc’s habitat and lifestyle. When the organism dies, its shell can become buried and preserved in sediments at the bottom of the lake or ocean (Figure 1d-f). Once preserved, these shells provide us with a valuable record of environmental conditions at the time that the organism lived. We can analyse the isotopes within the shells to build up a picture of environmental conditions in the past. -

FAO Fisheries & Aquaculture
Food and Agriculture Organization of the United Nations Fisheries and for a world without hunger Aquaculture Department Cultured Aquatic Species Information Programme Mercenaria mercenaria (Linnaeus, 1758) I. Identity V. Status And Trends a. Biological Features VI. Main Issues b. Images Gallery a. Responsible Aquaculture Practices II. Profile VII. References a. Historical Background a. Related Links b. Main Producer Countries c. Habitat And Biology III. Production a. Production Cycle b. Production Systems c. Diseases And Control Measures IV. Statistics a. Production Statistics b. Market And Trade Identity Mercenaria mercenaria Linnaeus, 1758 [Veneridae] FAO Names: En - Northern quahog(=Hard clam), Fr - Praire, Es - Chirla mercenaria Biological features Shell solid, equivalve; inequilateral, beaks in the front half of the shell; broadly oval in outline. Ligament a deeply inset, dark brown elliptical band, behind the beaks reaching half-way to the posterior margin. Lunule well defined, broad, heart-shaped. Escutcheon indistinct. Sculpture of concentric lines, raised here and there into ridges, and fine radiating lines. In young specimens the ridges are present all over the shell but in the adult they persist, after wear and tear, only near the anterior and posterior margins. Growth stages prominent. Both valves with three cardinal teeth; in addition there is present in each valve a rough tooth-like area behind the beaks and immediately below the ligament; this area has the appearance of a supplementary posterior cardinal tooth which has been broken off. No laterals. Pallial sinus not deep, triangular. Margin grenulate. Colour a dirty white, light varnish-brown, dull grey or grey-brown. Inside of shell white, sometimes deep violet about the adductor muscle scars. -

Mechanics Unlocks the Morphogenetic Puzzle of Interlocking Bivalved Shells
Mechanics unlocks the morphogenetic puzzle of interlocking bivalved shells Derek E. Moultona,1 , Alain Gorielya , and Regis´ Chiratb aMathematical Institute, University of Oxford, Oxford, OX2 6GG, United Kingdom; and bCNRS 5276, LGL-TPE (Le Laboratoire de Geologie´ de Lyon: Terre, Planetes,` Environnement), Universite´ Lyon 1, 69622 Villeurbanne Cedex, France Edited by Sean H. Rice, Texas Tech University, Lubbock, TX, and accepted by Editorial Board Member David Jablonski November 11, 2019 (received for review September 24, 2019) Brachiopods and mollusks are 2 shell-bearing phyla that diverged tal events causing shell injuries. Yet, in all cases the interlocking from a common shell-less ancestor more than 540 million years ago. of the 2 shell edges is tightly maintained. These observations Brachiopods and bivalve mollusks have also convergently evolved imply that the interlocking pattern emerges as the result of epi- a bivalved shell that displays an apparently mundane, yet strik- genetic interactions modulating the behavior of the secreting ing feature from a developmental point of view: When the shell mantle during shell development. is closed, the 2 valve edges meet each other in a commissure that Here, we provide a geometric and mechanical explanation forms a continuum with no gaps or overlaps despite the fact that for this morphological trait based on a detailed analysis of the each valve, secreted by 2 mantle lobes, may present antisymmet- shell geometry during growth and the physical interaction of the ric ornamental patterns of varying regularity and size. Interlock- shell-secreting soft mantle with both the rigid shell edge and ing is maintained throughout the entirety of development, even the opposing mantle lobe. -

Brief Glossary and Bibliography of Mollusks
A Brief Glossary of Molluscan Terms Compiled by Bruce Neville Bivalve. A member of the second most speciose class of Mollusca, generally bearing a shell of two valves, left and right, and lacking a radula. Commonly called clams, mussels, oysters, scallops, cockles, shipworms, etc. Formerly called pelecypods (class Pelecypoda). Cephalopoda. The third dominant class of Mollusca, generally without a true shell, though various internal hard structures may be present, highly specialized anatomically for mobility. Commonly called octopuses, squids, cuttles, nautiluses. Columella. The axis, real or imaginary, around and along which a gastropod shell grows. Dextral. Right-handed, with the aperture on the right when the spire is at the top. Most gastropods are dextral. Gastropod. A member of the largest class of Mollusca, often bearing a shell of one valve and an operculum. Commonly called snails, slugs, limpets, conchs, whelks, sea hares, nudibranchs, etc. Mantle. The organ that secretes the shell. Mollusk (or mollusc). A member of the second largest phylum of animals, generally with a non-segmented body divided into head, foot, and visceral regions; often bearing a shell secreted by a mantle; and having a radula. Operculum. A horny or calcareous pad that partially or completely closes the aperture of some gastropodsl. Periostracum. The proteinaceous layer covering the exterior of some mollusk shells. Protoconch. The larval shell of the veliger, often remains as the tip of the adult shell. Also called prodissoconch in bivlavles. Radula. A ribbon of teeth, unique to mollusks, used to procure food. Sinistral. Left-handed, with the aperture on the left when the spire is at the top. -

Proceedings of the Academy of Natural Sciences of Philadelphia
32 PROCEEDINGS OF THE ACADEMY OF [1887. ON NEW GENEEIC FOKMS OF CRETACEOUS MOLLUSCA AND THEIE RELATION TO OTHER FORMS. BY CHARLES A. WHITE. Published by permission of the Director of the United States Geological Survey. The type species of the three generic forms which are described in this article ^ belong to the collections of Cretaceous fossils from Texas, which I am now preparing for publication in one of the memoirs of the U. S. Geological Survey. In their generic charac- teristics all three of them appear to be respectively identical with certain forms which have long been known, but which have been referred to other genera by different authors. The features which I now present as having generic value seem to have been overlooked by those authors, or, so far as they were observed, they were treated as specific characters. Two of these forms belong to the section Melininse of the family Aviculidse. The other is referred to the Crassatellidse, but it departs considerably from the typical section of that family. CRASSATELLIDJE. Genus STEARNSIA (gen. nov.). Shell compressed, subtrihedral or subcircular in marginal out- line; beaks small, closely approximate, prominent by reason of the abrupt sloping away of both the antero-and postero-dorsal borders; lunule and escutcheon both well defined and flattened or excavated; hinge strong, consisting of both cardinal and lateral teeth; cardinal teeth two in the left valve and three in the right; both posterior and anterior lateral teeth long and slender; posterior laterals two in the right valve and one in the left; anterior laterals two in the left valve and one in the right. -

Three Alien Molluscs from Iskenderun Bay (SE Turkey)
Aquatic Invasions (2006) Volume 1, Issue 2: 76-79 DOI 10.3391/ai.2006.1.2.4 © 2006 The Author(s) Journal compilation © 2006 REABIC (http://www.reabic.net) This is an Open Access article Research article Three alien molluscs from Iskenderun Bay (SE Turkey) Doğan Çeviker1 and Serhat Albayrak2* 1Itri Sokak No:2 34349 Balmumcu-Istanbul, Turkey E-mail: [email protected] 2Istanbul University, Faculty of Science, Department of Biology 34118 Vezneciler-Istanbul, Turkey E-mail: [email protected] *Corresponding author Received 26 April 2006; accepted in revised form 4 May 2006 Abstract This study reports the presence of three alien molluscs from Iskenderun Bay (SE Turkey). Amathina tricarinata (Linnaeus, 1767) and Petricola hemprichi Issel, 1869 have prior records from other regions of Mediterranean, but, Cardites akabana (Sturany, 1899) first recorded in this paper. Since all of them are present in the Red Sea or Suez Canal, they can be considered as Lessepsian immigrants. Key words: Mollusca, alien species, Mediterranean, Turkey Introduction that 88 % of the exotic molluscs are Lessepsian immigrants in the eastern Mediterranean (Galil The Mediterranean Sea hosts about 8500 species and Zenetos 2002). Detailed data about these species of macroscopic animals. This rich biodiversity, are available on the Internet (www.ciesm.org/atlas). representing 8-9 % of total species number of the Either Lessepsian or non-Lessepsian, many world’s seas, comprises temperate and sub- new non-indigenous species continue to enter the tropical elements together with endemic and Mediterranean. alien species (Zenetos et al. 2002). The eastern Mediterranean is most vulnerable The introduction of alien species (also known to invasion and should be continuously as exotic, introduced or non-native species) into monitored. -

The Impact of Mollusks on Water Quality
The Impact of Mollusks on Water Quality SYNOPSIS VARIATIONS 1 TO 9 Students will determine how a filter-feeding TEACHER PARTNER SCIENTIST PARTNER mollusk, such as an oyster, clam, or mussel, re- Blair Buck Roger Newell sponds to changes in its immediate environ- Episcopal High School Horn Point Laboratory ment. Students will design experiments to test 1200 Quaker Lane University of MD Center hypotheses about the effects of environmental Alexandria, VA 22302 for Environmental Science variables on filter feeding and identify anatomi- PO Box 775 cal structures of a mollusk. Cambridge, MD 21613 CORE EXPERIMENT TEACHER SCIENTIST PARTNER Joe Carter APPROPRIATE BIOLOGY LEVEL PARTNERS University of North Introductory, advanced, ecology Kathy Dunnavant and Cyndi Louden Carolina at Chapel Hill Cary High School Geology Department Walnut Street CB3315 Cary, NC 27511 Mitchell Hall Chapel Hill, NC 27599-3315 Directions for Teachers Note to Teachers: Information is given for the Core Experiment below. Additional information needed for each variation of the Core Experiment may be found beginning LENGTH OF LAB on page 215. For a specific variation, check the At-A-Glance Map. A suggested time allotment is as follows: Day 1 (45 minutes) GETTING READY • Demonstrate filtration and design See sidebars for additional information regarding preparation of the lab. experiment. Day 2 (45 minutes) OBJECTIVES FOR CORE EXPERIMENT • Set up experiment. Place animals into treatment aquaria. At the end of this lab, students will be able to: Day 3 (20 minutes) • Describe the ecological role of filter-feeding mollusks in fresh and/or saltwater • Move animals from treatment tanks ecosystems. to collection containers. -

Structure and Function of the Digestive System in Molluscs
Cell and Tissue Research (2019) 377:475–503 https://doi.org/10.1007/s00441-019-03085-9 REVIEW Structure and function of the digestive system in molluscs Alexandre Lobo-da-Cunha1,2 Received: 21 February 2019 /Accepted: 26 July 2019 /Published online: 2 September 2019 # Springer-Verlag GmbH Germany, part of Springer Nature 2019 Abstract The phylum Mollusca is one of the largest and more diversified among metazoan phyla, comprising many thousand species living in ocean, freshwater and terrestrial ecosystems. Mollusc-feeding biology is highly diverse, including omnivorous grazers, herbivores, carnivorous scavengers and predators, and even some parasitic species. Consequently, their digestive system presents many adaptive variations. The digestive tract starting in the mouth consists of the buccal cavity, oesophagus, stomach and intestine ending in the anus. Several types of glands are associated, namely, oral and salivary glands, oesophageal glands, digestive gland and, in some cases, anal glands. The digestive gland is the largest and more important for digestion and nutrient absorption. The digestive system of each of the eight extant molluscan classes is reviewed, highlighting the most recent data available on histological, ultrastructural and functional aspects of tissues and cells involved in nutrient absorption, intracellular and extracellular digestion, with emphasis on glandular tissues. Keywords Digestive tract . Digestive gland . Salivary glands . Mollusca . Ultrastructure Introduction and visceral mass. The visceral mass is dorsally covered by the mantle tissues that frequently extend outwards to create a The phylum Mollusca is considered the second largest among flap around the body forming a space in between known as metazoans, surpassed only by the arthropods in a number of pallial or mantle cavity.