Intrinsic Hand Muscles of the Japanese Monkey, Macaca Fuscata
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pronator Syndrome: Clinical and Electrophysiological Features in Seven Cases
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.39.5.461 on 1 May 1976. Downloaded from Journal ofNeurology, Neurosurgery, and Psychiatry, 1976, 39, 461-464 Pronator syndrome: clinical and electrophysiological features in seven cases HAROLD H. MORRIS AND BRUCE H. PETERS From the Department ofNeurology, University of Texas Medical Branch, Galveston, Texas, USA SYNOPSIS The clinical and electrophysiological picture of seven patients with the pronator syndrome is contrasted with other causes ofmedian nerve neuropathy. In general, these patients have tenderness over the pronator teres and weakness of flexor pollicis longus as well as abductor pollicis brevis. Conduction velocity of the median nerve in the proximal forearm is usually slow but the distal latency and sensory nerve action potential at the wrist are normal. Injection of corticosteroids into the pronator teres has produced relief of symptoms in a majority of patients. Protected by copyright. In the majority of isolated median nerve dys- period 101 cases of the carpal tunnel syndrome functions the carpal tunnel syndrome is appropri- and the seven cases of the pronator syndrome ately first suspected. The median nerve can also reported here were identified. Median nerve be entrapped in the forearm giving rise to a conduction velocity determinations were made on similar picture and an erroneous diagnosis. all of these patients. The purpose of this report is to draw full attention to the pronator syndrome and to the REPORT OF CASES features which allow it to be distinguished from Table 1 provides clinical details of seven cases of the median nerve entrapment at other sites. -

Wrist and Hand Examina[On
Wrist and Hand Examinaon Daniel Lueders, MD Assistant Professor Physical Medicine and Rehabilitaon Objecves • Understand the osseous, ligamentous, tendinous, and neural anatomy of the wrist and hand • Outline palpable superficial landmarks in the wrist and hand • Outline evaluaon of and differen.aon between nerves to the wrist and hand • Describe special tes.ng of wrist and hand Wrist Anatomy • Radius • Ulna • Carpal bones Wrist Anatomy • Radius • Ulna • Carpal bones Wrist Anatomy • Radius • Ulna • Carpal bones Wrist Anatomy • Radius • Ulna • Carpal bones Inspec.on • Ecchymosis • Erythema • Deformity • Laceraon Inspec.on • Common Finger Deformies • Swan Neck Deformity • Boutonniere Deformity • Hypertrophic nodules • Heberden’s, Bouchard’s Inspec.on • Swan Neck Deformity • PIP hyperextension, DIP flexion • Pathology is at PIP joint • Insufficiency of volar/palmar plate and suppor.ng structures • Distally, the FDP tendon .ghtens from PIP extension causing secondary DIP flexion • Alternavely, extensor tendon rupture produces similar deformity Inspec.on • Boutonniere Deformity • PIP flexion, DIP hyperextension • Pathology is at PIP joint • Commonly occurs from insufficiency of dorsal and lateral suppor.ng structures at PIP joint • Lateral bands migrate volar/palmar, creang increased flexion moment • Results in PIP “buTon hole” effect dorsally Inspec.on • Nodules • Osteoarthri.c • Hypertrophic changes of OA • PIP - Bouchard’s nodule • DIP - Heberden’s nodule • Rheumatoid Arthri.s • MCP joints affected most • Distal radioulnar joint can also be affected -

Nerve Transfer Techniques in Injuries from the Upper Limb
THIEME Update Article | Artículo de Actualización 57 Nerve Transfer Techniques in Injuries from the Upper Limb Técnicas de transferencia nerviosa en lesiones del miembro superior Francisco Martínez Martínez1 B. Ñíguez Sevilla2 J. García García2 A. García López3 1 FEA (field medcial expert), Orthopaedics and Traumatology Surgery, Address for correspondence Francisco Martínez Martínez, C/ Canovas Hospital Clínico Universitario Virgen de la Arrixaca. Murcia, Spain del Castillo n°7- 4°a. 30003-Murcia, Spain 2 Resident, Orthopaedics and Traumatology Surgery, Hospital Clínico (e-mail: [email protected]). Universitario Virgen de la Arrixaca, Murcia, Spain 3 FEA (field medcial expert), Orthopaedics and Traumatology Surgery, Hospital General de Alicante, Alicante, Spain Rev Iberam Cir Mano 2017;45:57–67. Abstract Proximal nerve injuries from the upper limb or the braquial plexus are associated with a poor prognosis, even with prompt repair. In the last few decades an increase in nerve transfer techniques has occurred, by which a denervated peripheral nerve is reinner- vated by a healthy donor nerve. Nerve transfers are indicated in proximal brachial plexus injuries where grafting is not possible or in proximal injuries of peripheral nerves with long reinnervation distances. Nerve transfers represent a revolution in peripheral nerve surgery and offer the potential for superior functional recovery in severe nerve injuries. In complete brachial plexus injuries, there are being studied the existence of nerve roots (intraplexual transfers). If they do not exist, the transference of nerves out of the plexus are used Keywords (extraplexual transfers) as the spinal accessory nerve, the phrenic nerve, the intercostal ► nerve transfers nerves, etc. ► brachial plexus In this update paper, the different motor intra and extraplexual nerve transfer ► nerve injury techniques are going to be reviewed. -

The Muscles That Act on the Upper Limb Fall Into Four Groups
MUSCLES OF THE APPENDICULAR SKELETON UPPER LIMB The muscles that act on the upper limb fall into four groups: those that stabilize the pectoral girdle, those that move the arm, those that move the forearm, and those that move the wrist, hand, and fingers. Muscles Stabilizing Pectoral Girdle (Marieb / Hoehn – Chapter 10; Pgs. 346 – 349; Figure 1) MUSCLE: ORIGIN: INSERTION: INNERVATION: ACTION: ANTERIOR THORAX: anterior surface coracoid process protracts & depresses Pectoralis minor* pectoral nerves of ribs 3 – 5 of scapula scapula medial border rotates scapula Serratus anterior* ribs 1 – 8 long thoracic nerve of scapula laterally inferior surface stabilizes / depresses Subclavius* rib 1 --------------- of clavicle pectoral girdle POSTERIOR THORAX: occipital bone / acromion / spine of stabilizes / elevates / accessory nerve Trapezius* spinous processes scapula; lateral third retracts / rotates (cranial nerve XI) of C7 – T12 of clavicle scapula transverse processes upper medial border elevates / adducts Levator scapulae* dorsal scapular nerve of C1 – C4 of scapula scapula Rhomboids* spinous processes medial border adducts / rotates dorsal scapular nerve (major / minor) of C7 – T5 of scapula scapula * Need to be familiar with on both ADAM and the human cadaver Figure 1: Muscles stabilizing pectoral girdle, posterior and anterior views 2 BI 334 – Advanced Human Anatomy and Physiology Western Oregon University Muscles Moving Arm (Marieb / Hoehn – Chapter 10; Pgs. 350 – 352; Figure 2) MUSCLE: ORIGIN: INSERTION: INNERVATION: ACTION: intertubercular -

Ulnar Nerve Passing Through Triceps Muscle –Rare Variation
IOSR Journal of Dental and Medical Sciences (IOSR-JDMS) e-ISSN: 2279-0853, p-ISSN: 2279-0861. Volume 12, Issue 6 (Nov.- Dec. 2013), PP 61-62 www.iosrjournals.org Ulnar Nerve Passing Through Triceps Muscle –Rare Variation Chandrika G Teli1, Nilesh N. Kate2, H. S. Kadlimatti 3 1(Department of Anatomy, ESIC Medical College Gulbarga/ Rajiv Gandhi university of health sciences Karnataka, India) 2(Department of Physiology, ESIC Medical College Gulbarga / Rajiv Gandhi university of health sciences Karnataka, India) 3(Department of Anatomy, ESIC Medical College Gulbarga / Rajiv Gandhi university of health sciences Karnataka, India) Abstract : During routine dissection for undergraduate students, right upper limb of 45 year old male showed variation in the course of ulnar nerve. The ulnar nerve arose as a continuation of medial cord, descended down along medial side of axillary and brachial artery. As the nerve travelled upper third of arm, it pierced the medial head belly of triceps muscle, passing through it for 4-5 cm ,emerged out of it to reach near the medial epicondyle. Then the nerve passed behind the medial epicondyle to follow its normal course and distribution. This kind of variation is not been described in the literature. Keywords: lunar nerve, variation in course, entrapment neuropathy. I. Introduction The ulnar nerve (C7, 8, T1) is formed from medial cord of the brachial plexus. It lies medial to axillary and brachial artery as far as middle of humerus, and then pierces the medial inter muscular septum to descend on the anterior face of triceps. It passes behind the medial epicondyle to enter the forearm. -
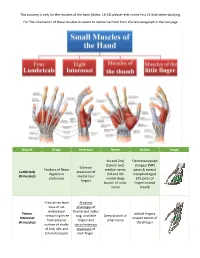
This Sumarry Is Only for the Muscles of the Hand (Slides: 14-33) Please Refer to the First 13 Slide When Studying
This sumarry is only for the muscles of the hand (slides: 14-33) please refer to the first 13 slide when studying. For The innervation of these muscles its easier to memorize them from the last paragraph in the last page Muscle Origin Insertion Nerve Action Image 1st and 2nd, Flexmetacarpoph (lateral two) alangeal (MP) Extensor Tendons of flexor median nerve; joints & extend Lumbricals expansion of Digitorum 3rd and 4th interphalangeal (4 muscles) medial four profundus medial deep (IP) joints of fingers branch of ulnar fingers except nerve thumb -First arises from Proximal base of 1st phalanges of metacarpal thumb and index, Palmar adduct fingers - remaining three ring, and little Deep branch of Interossei toward center of from anterior fingers and ulnar nerve (4 muscles) third finger surface of shafts dorsal extensor of 2nd, 4th, and expansion of 5th metacarpals each finger . Proximal phalanges of index, middle, and ring fingers Contiguous sides abduct fingers Dorsal Interossei and dorsal Deep branch of of shafts of from center of (4 muscles) extensor ulnar nerve metacarpal bones third finger expansion (1st:index\ 2nd,3rd:middle \ 4th:ring) Both palmar and dorsal: -Flex metacarpophalangeal joints -Extend interphalangeal joints Simultaneous flexion at the metacarpophalangeal joints and extension at the interphalangeal joints of a digit are essential for the fine movements of writing, drawing, threading a needle, etc. The Lumbricals and interossei have long been accepted as not only primary agents in flexing the metacarpophalangeal joints -

Morphological Study of Palmaris Longus Muscle
International INTERNATIONAL ARCHIVES OF MEDICINE 2017 Medical Society SECTION: HUMAN ANATOMY Vol. 10 No. 215 http://imedicalsociety.org ISSN: 1755-7682 doi: 10.3823/2485 Humberto Ferreira Morphological Study of Palmaris Arquez1 Longus Muscle ORIGINAL 1 University of Cartagena. University St. Thomas. Professor Human Morphology, Medicine Program, University of Pamplona. Morphology Laboratory Abstract Coordinator, University of Pamplona. Background: The palmaris longus is one of the most variable muscle Contact information: in the human body, this variations are important not only for the ana- tomist but also radiologist, orthopaedic, plastic surgeons, clinicians, Humberto Ferreira Arquez. therapists. In view of this significance is performed this study with Address: University Campus. Kilometer the purpose to determine the morphological variations of palmaris 1. Via Bucaramanga. Norte de Santander, longus muscle. Colombia. Suramérica. Tel: 75685667-3124379606. Methods and Findings: A total of 17 cadavers with different age groups were used for this study. The upper limbs region (34 [email protected] sides) were dissected carefully and photographed in the Morphology Laboratory at the University of Pamplona. Of the 34 limbs studied, 30 showed normal morphology of the palmaris longus muscle (PL) (88.2%); PL was absent in 3 subjects (8.85% of all examined fo- rearm). Unilateral absence was found in 1 male subject (2.95% of all examined forearm); bilateral agenesis was found in 2 female subjects (5.9% of all examined forearm). Duplicated palmaris longus muscle was found in 1 male subject (2.95 % of all examined forearm). The palmaris longus muscle was innervated by branches of the median nerve. The accessory palmaris longus muscle was supplied by the deep branch of the ulnar nerve. -

ISPUB.COM Volume 9 Number 2
The Internet Journal of Surgery ISPUB.COM Volume 9 Number 2 An Anomalous Palmaris Brevis Muscle and its Clinical Implications S Das, S Paul Citation S Das, S Paul. An Anomalous Palmaris Brevis Muscle and its Clinical Implications. The Internet Journal of Surgery. 2006 Volume 9 Number 2. Abstract The palmaris brevis (PB) muscle is a small, thin, subcutaneous muscle of the palm. The palmaris brevis muscle is important muscle contributing to the grip of the hand. In the present case, during routine cadaveric dissection, we detected a PB muscle which was not subcutaneous but existed as a muscle deep to the dermis of the skin of the palm. The PB muscle stretched as a horizontal band over the hypothenar muscles of the palm. The anomalous location of the PB may result in compression of neurovascular structures on the ulnar side of the palm. The awareness of such anomalous muscle which is not subcutaneous, may be clinical important for surgeons operating on the hand and clinicians diagnosing compressive symptoms. INTRODUCTION Figure 1 The PB muscle is a thin subcutaneous muscle. The PB arises Figure 1: Photograph of dissected right palm showing from the flexor retinaculum and the medial border of the central part of the palmar aponeurosis and attached to the dermis of the skin on the ulnar side of the palm. 1 The PB is innervated by the ulnar nerve and the main action of the muscle makes the palmar grip more secure. 1 The clinical importance of the muscle lies in the fact, that it lies above the ulnar artery and the ulnar nerve in the palm. -

Ulnar Nerve Contribution in the Innervation of the Triceps Brachii Muscle Ulnar Nerve to the Triceps Brachii
ORIGINAL COMMUNICATION Anatomy Journal of Africa. 2017. Vol 6 (1): 834 – 839. ULNAR NERVE CONTRIBUTION IN THE INNERVATION OF THE TRICEPS BRACHII MUSCLE ULNAR NERVE TO THE TRICEPS BRACHII Silva DLR, Barros MP, Freire TGS, Firmino Júnior L, Almeida Filho WRB, Correia Cadeira JSL, Silva NO CorresPondence to Diêgo Lucas Ramos e Silva Rua Prof. Virgilio Guedes, 1391 Ponta Grossa, 57014-220 Maceió, AL. Email: [email protected] ABSTRACT The ulnar nerve is considered the thickest terminal branch of the medial cord in the brachial Plexus and most authors does not mention the possibility of this nerve emitting branches to the arm. However, some studies rePorted that the ulnar nerve could suPPly the medial head of triceps brachii muscle. The main objective in this study was identifying the Presence of ulnar nerve branches in tricePs brachii muscle. Sixty uPPer limbs of adult Brazilian corpses of both sexes were used. The estimated age was between 25 and 80 years old. Every studied piece had the nerves and their branches quantified and measured with a manual mechanic caliper. The branches were photographed and had the data registered in individual files. Were found ulnar nerve branches for all the heads of tricePs brachii muscle: 1 branch (9,1%) to lateral head, 2 branches (18,1%) to long head and 8 branches (72,7%) to medial head. Thus, we can conclude that the contribution of ulnar nerve to tricePs brachii muscle constitutes an imPortant anatomical variation. Key words: Ulnar nerve; Triceps brachii muscle; Innervation. INTRODUCTION The ulnar nerve is the terminal branch of the communication with the radial nerve (Bekler et medial cord of the brachial Plexus, receiving C8 al, 2009). -

Pain in the Thenar Eminence: a Rare Case of Atypical Angina
782 BRITISH MEDICAL JOURNAL VOLUME 281 20 SEPTEMBER 1980 Comment Treatment with propranolol 80 mg three times daily produced some improvement, but she continued to have spontaneous attacks of pain. Deaths from rhesus haemolytic disease of the newborn have been Verapamil 120 mg three times daily produced further improvement, but Br Med J: first published as 10.1136/bmj.281.6243.782 on 20 September 1980. Downloaded from declining for many years, but since the introduction of anti-D the fall she continued to have pain while resting so a bypass graft was considered. has been much steeper.5 The patient refused surgery, and the dose of verapamil was increased to It is not possible to draw definite conclusions from data based on 120 mg four times daily. Over the next month her symptoms resolved, only two years, but trends that would be expected if the prophylaxis with an increase in treadmill exercise time to 5-2 minutes and only one was successful (assuming treatment of rhesus haemolytic disease of episode of notable ST depression in 24 hours of continuous monitoring. the newborn remains substantially unchanged) may be assessed. Given the natural history of worsening of the disease in consecutive 2mm] babies, the reduction in deaths is more likely to be seen initially in the livebom babies. This is because mothers of stillborn babies will usually have been immunised for longer than mothers of liveborn CC5 + + babies-and many of them will still be in categories 1 and 2. On the other hand, the numbers in categories 3 and 4 should remain approximately constant since in neither group is prophylactic anti-D administered. -

Upper and Lower Extremity Nerve Conduction Studies Kelly G
2019 Upper and Lower Extremity Nerve Conduction Studies Kelly G. Gwathmey October 18, 2019 Virginia Commonwealth University 2019 Financial Disclosure I have received speaking and consulting honoraria from Alexion Pharmaceuticals. 2019 Warning Videotaping or taking pictures of the slides associated with this presentation is prohibited. The information on the slides is copyrighted and cannot be used without permission and author attribution. 2019 Outline for Today’s talk • Upper extremity nerve conduction studies o Median nerve o Ulnar nerve o Radial nerve o Median comparison studies o Medial antebrachial cutaneous nerve o Lateral antebrachial cutaneous nerve • Lower extremity nerve conduction studies o Fibular nerve o Tibial nerve o Sural nerve o Femoral nerve • Saphenous • Lateral femoral cutaneous • Phrenic nerve • Facial nerve • Anomalous Innervations 2019 Median nerve anatomy • Median nerve is formed by a combination of: o Lateral cord (C6-7) supplies the sensory fibers to the thumb, index, middle finger, proximal median forearm, and thenar eminence. o Medial cord (C8-T1) provides motor fibers to the distal forearm and hand. • The median nerve innervates the pronator teres, then gives branches to the flexor carpi radialis, flexor digitorum superficialis, and palmaris longus. • Anterior Interosseus Nerve (AIN)- innervates the flexor pollicis longus, flexor digitorum profundus (FDP) (digits 2 and 3), and pronator quadratus. Preston, David C., MD; Shapiro, Barbara E., MD, PhD. Published January 1, 2013. Pages 267-288. © 2013. 2019 Median nerve anatomy • Proximal to the wrist- the palmar cutaneous sensory branch (sensation over the thenar eminence) • Through the carpal tunnel- Motor division goes to first and second lumbricals o Recurrent thenar motor branch the thenar eminence (opponens, abductor pollicis brevis, and superficial head of flexor pollicis brevis) • Sensory branch that goes through the carpal tunnel supplies the medial thumb, index finger, middle finger and lateral half of the ring finger. -

Carpal Tunnel Syndrome • Link to This Article Online for CPD/CME Credits Scott D Middleton,1 Raymond E Anakwe2
EDUCATION CLINICAL REVIEW Carpal tunnel syndrome • Link to this article online for CPD/CME credits Scott D Middleton,1 Raymond E Anakwe2 1Department of Trauma and Orthopaedic Surgery, Royal Carpal tunnel syndrome is the most commonly diagnosed Infirmary of Edinburgh, Edinburgh, SOURCES AND SELECTION CRITERIA compression neuropathy of the upper limb. Patients may UK We searched PubMed, the Cochrane Library, and the 2Department of Trauma and present to general practitioners, physiotherapists, hand Cumulative Index to Nursing and Allied Health Literature Orthopaedic Surgery, St Mary’s therapists, or surgeons with a variety of symptoms. Sev- to identify source material for this review. We examined Hospital, Imperial College NHS eral studies have examined the epidemiology, diagno- available evidence published in the English language for Trust, London W1 2NY, UK Correspondence to: R E Anakwe sis, and treatment of carpal tunnel syndrome. We review the diagnosis and treatment of carpal tunnel syndrome. [email protected] these resources to provide an evidence based guide to The search terms used were “carpal tunnel”, “carpal Cite this as: BMJ 2014;349:g6437 the diagnosis and treatment of carpal tunnel syndrome. tunnel syndrome”, “tingling fingers”, “median nerve doi: 10.1136/bmj.g6437 compression”, and “compression neuropathy”. There were no well conducted large randomised trials. We selected and What is carpal tunnel syndrome and who gets it? examined smaller randomised trials as well as case series, Carpal tunnel syndrome encompasses a collection of cohort studies, and observational reports where these symptoms: patients often mention altered sensation or provided the only evidence. pain in the hand, wrist, or forearm.