SPECIES Isocoma Menziesii (Hooker & Arnnott) G
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rare Native Vascular Plants S
RARE NATIVE VASCULAR PLANTS OF THE SOUTHERN OKANAGAN GRASSLANDS PROTECTED AREA FIELD REPORT " Douglas Ecological Consultants 2004 by George W. Douglas and Shyanne 1. Smith Douglas Ecological Consultants Ltd. August 12,2004 Funding Provided by The Nature Trust of British Columbia through the BrinklMcLean Grassland Conservation Fund OVERVIEW The Southern Okanagan Grasslands Protected Area was established in 2001 and encompasses about 4,481 ha of native grasslands in the southern Okanagan and southern Similkameen valleys of British Columbia. In the summer of2004, The Nature Trust, through the Brink/McLean Grassland Conservation Fund, provided Douglas Ecological Consultants Ltd. with funds to conduct a two-day inventory in the protected area for rare native vascular plants. The rare plants searched for during the inventory are those appearing on the British Columbia Conservation Data Centre's (BC CDC) Red/Blue lists (Douglas et a.12002). The study area has not been well inventoried and current knowledge, with respect to vascular plants, is minimal. METHODS Due to the brief inventory period, only a small part of the Southern Okanagan Grasslands Protected Area was examined. The study area consisted of a corridor about five km long and about 200 m wide through the Protected Area (Figure 1). This corridor followed the main, east-west dirt tract through the area, thus minimizing travel time and maximizing search time. The two-day search methodology consisted of examining various micro-habitats along the corridor likely to contain different species of rare plants. Once a rare plant population was located, a number of population attributes were recorded. These attributes included: GPS location, elevation, slope, aspect, habitat, plant associates in habitat, size of habitat, quality of the habitat, number of rare plant(s) present and quality of the element plant occurrence!. -

California Vegetation Map in Support of the DRECP
CALIFORNIA VEGETATION MAP IN SUPPORT OF THE DESERT RENEWABLE ENERGY CONSERVATION PLAN (2014-2016 ADDITIONS) John Menke, Edward Reyes, Anne Hepburn, Deborah Johnson, and Janet Reyes Aerial Information Systems, Inc. Prepared for the California Department of Fish and Wildlife Renewable Energy Program and the California Energy Commission Final Report May 2016 Prepared by: Primary Authors John Menke Edward Reyes Anne Hepburn Deborah Johnson Janet Reyes Report Graphics Ben Johnson Cover Page Photo Credits: Joshua Tree: John Fulton Blue Palo Verde: Ed Reyes Mojave Yucca: John Fulton Kingston Range, Pinyon: Arin Glass Aerial Information Systems, Inc. 112 First Street Redlands, CA 92373 (909) 793-9493 [email protected] in collaboration with California Department of Fish and Wildlife Vegetation Classification and Mapping Program 1807 13th Street, Suite 202 Sacramento, CA 95811 and California Native Plant Society 2707 K Street, Suite 1 Sacramento, CA 95816 i ACKNOWLEDGEMENTS Funding for this project was provided by: California Energy Commission US Bureau of Land Management California Wildlife Conservation Board California Department of Fish and Wildlife Personnel involved in developing the methodology and implementing this project included: Aerial Information Systems: Lisa Cotterman, Mark Fox, John Fulton, Arin Glass, Anne Hepburn, Ben Johnson, Debbie Johnson, John Menke, Lisa Morse, Mike Nelson, Ed Reyes, Janet Reyes, Patrick Yiu California Department of Fish and Wildlife: Diana Hickson, Todd Keeler‐Wolf, Anne Klein, Aicha Ougzin, Rosalie Yacoub California -

Artemisia Californica Less
I. SPECIES Artemisia californica Less. [Updated 2017] NRCS CODE: Subtribe: Artemisiinae ARCA11 Tribe: Anthemideae (FEIS CODE: Family: Asteraceae ARCAL) Order: Asterales Subclass: Asteridae Class: Magnoliopsida flowering heads spring growth seedling, March 2009 juvenile plant photos A. Montalvo flowering plant, November 2005 mature plant with flower buds August 2010 A. Subspecific taxa None. Artemisia californica Less. var. insularis (Rydb.) Munz is now recognized as Artemisia nesiotica P.H. Raven (Jepson eFlora 2017). B. Synonyms Artemisia abrotanoides Nuttall; A. fischeriana Besser; A. foliosa Nuttall; Crossostephium californicum (Lessing) Rydberg (FNA 2017). C. Common name California sagebrush. The common name refers to its strong, sage-like aroma and endemism to California and Baja California. Other names include: coastal sage, coast sage, coast sagebrush (Painter 2016). D. Taxonomic relationships The FNA (2017) places this species in subgenus Artemisia . The molecular phylogeny of the genus has improved the understanding of relationships among the many species of Artemisia and has, at times, placed the species in subgenus Tridentadae; morphology of the inflorescences and flowers alone does not place this species with its closest relatives (Watson et al. 2002). The detailed phylogeny is not completely resolved (Hayat et al. 2009). E. Related taxa in region There are 18 species and a total of 31 taxa (including infrataxa) of Artemisia in southern California, all of which differ clearly from A. californica in habitat affinity, structure, or both (Munz 1974, Jepson eFlora 2017). Within subgenus Artemisia (as per FNA 2017), A. nesiotica from the Channel Islands is the most similar and was once considered part of A. californica ; it can be distinguished by its wider leaves with flat leaf margins (not rolled under). -

2014 Plant Species List
Acanthaceae Hygrophila Occasional lacustris Acanthaceae Justicia ovata Uncommon Acanthaceae Ruellia humilis Common Acanthaceae Ruellia nudiflora s.n. Uncommon Acanthaceae Ruellia Occasional pedunculata Aceraceae Acer rubrum Occasional Agavaceae Yucca louisianica Uncommon Aiozaceae Molluga Occasional verticillata Alismataceae Echinodorus Occasional cordifolius Alismataceae Sagittaria Rare papillosa Alismataceae Sagittaria 156 Uncommon platyphylla Alliaceae Allium Occasional canadense var. canadense Alliaceae Allium Occasional canadense var. mobilense Alliaceae Allium 96, Uncommon drummondii 124 (Keith 96, 124) Amaranthaceae Alternanthera Common philoxeroides Amaryllidaceae Hymenocallis Uncommon liriosome Anacardiaceae Rhus aromatica Uncommon Anacardiaceae Rhus copallinum Occasional Anacardiaceae Toxicodendron Frequent radicans Apiaceae Bifora americana Common Apiaceae Centella erecta Uncommon Apiaceae Chaerophyllum Uncommon tainturieri Apiaceae Cicuta Uncommon maculatum Apiaceae Cynosciadium Uncommon digitatum Apiaceae Eryngium Common yuccifolium Apiaceae Hydrocotyle Occasional verticillata Apiaceae Polytaenia Frequent texana Apiaceae Ptilimnium Common capillaceum Apiaceae Ptilimnium Common nuttallii Apiaceae Spermolepsis Common inermis Apiaceae Torilis arvensis Occasional Apocynaceae Apocynum Occasional cannibinum Apocynaceae Nerium oleander Rare Apocynaceae Trachelospermu Occasional m difforme Aquifoliaceae Ilex decidua Common Aquifoliaceae Ilex opaca Common Aquifoliaceae Ilex vomitoria Abundant Araceae Arisaema Rare dracontium Araceae -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Notes on Variation and Geography in Rayjacksonia Phyllocephala (Asteraceae: Astereae)
Nesom, G.L., D.J. Rosen, and S.K. Lawrence. 2013. Notes on variation and geography in Rayjacksonia phyllocephala (Asteraceae: Astereae). Phytoneuron 2013-53: 1–15. Published 12 August 2013. ISSN 2153 733X NOTES ON VARIATION AND GEOGRAPHY IN RAYJACKSONIA PHYLLOCEPHALA (ASTERACEAE: ASTEREAE) GUY L. NESOM 2925 Hartwood Drive Fort Worth, Texas 76109 [email protected] DAVID J. ROSEN Department of Biology Lee College Baytown, Texas 77522-0818 [email protected] SHIRON K. LAWRENCE Department of Biology Lee College Baytown, Texas 77522-0818 ABSTRACT Inflorescences of Rayjacksonia phyllocephala in the disjunct Florida population system are characterized by heads on peduncles with leaves mostly reduced to linear bracts; heads in inflorescences of the Mexico-Texas-Louisiana system are immediately subtended by relatively unreduced leaves. The difference is consistent and justifies recognition of the Florida system as R. phyllocephala var. megacephala (Nash) D.B. Ward. Scattered waifs between the two systems are identified here as one or the other variety, directly implying their area of origin. In the eastern range of var. phyllocephala , at least from Brazoria County, Texas, eastward about 300 miles to central Louisiana, leaf margins vary from entire to deeply toothed-spinulose. In contrast, margins are invariably toothed-spinulose in var. megacephala as well as in the rest of southeastern Texas (from Brazoria County southwest) into Tamaulipas, Mexico. In some of the populations with variable leaf margins, 70-95% of the individuals have entire to mostly entire margins. KEY WORDS : Rayjacksonia , morphological variation, leaf margins, disjunction, waifs Rayjacksonia phyllocephala (DC.) Hartman & Lane (Gulf Coast camphor-daisy) is an abundant and conspicuous species of the shore vegetation around the Gulf of Mexico. -

IP Athos Renewable Energy Project, Plan of Development, Appendix D.2
APPENDIX D.2 Plant Survey Memorandum Athos Memo Report To: Aspen Environmental Group From: Lehong Chow, Ironwood Consulting, Inc. Date: April 3, 2019 Re: Athos Supplemental Spring 2019 Botanical Surveys This memo report presents the methods and results for supplemental botanical surveys conducted for the Athos Solar Energy Project in March 2019 and supplements the Biological Resources Technical Report (BRTR; Ironwood 2019) which reported on field surveys conducted in 2018. BACKGROUND Botanical surveys were previously conducted in the spring and fall of 2018 for the entirety of the project site for the Athos Solar Energy Project (Athos). However, due to insufficient rain, many plant species did not germinate for proper identification during 2018 spring surveys. Fall surveys in 2018 were conducted only on a reconnaissance-level due to low levels of rain. Regional winter rainfall from the two nearest weather stations showed rainfall averaging at 0.1 inches during botanical surveys conducted in 2018 (Ironwood, 2019). In addition, gen-tie alignments have changed slightly and alternatives, access roads and spur roads have been added. PURPOSE The purpose of this survey was to survey all new additions and re-survey areas of interest including public lands (limited to portions of the gen-tie segments), parcels supporting native vegetation and habitat, and windblown sandy areas where sensitive plant species may occur. The private land parcels in current or former agricultural use were not surveyed (parcel groups A, B, C, E, and part of G). METHODS Survey Areas: The area surveyed for biological resources included the entirety of gen-tie routes (including alternates), spur roads, access roads on public land, parcels supporting native vegetation (parcel groups D and F), and areas covered by windblown sand where sensitive species may occur (portion of parcel group G). -

Mcgrath State Beach Plants 2/14/2005 7:53 PM Vascular Plants of Mcgrath State Beach, Ventura County, California by David L
Vascular Plants of McGrath State Beach, Ventura County, California By David L. Magney Scientific Name Common Name Habit Family Abronia maritima Red Sand-verbena PH Nyctaginaceae Abronia umbellata Beach Sand-verbena PH Nyctaginaceae Allenrolfea occidentalis Iodinebush S Chenopodiaceae Amaranthus albus * Prostrate Pigweed AH Amaranthaceae Amblyopappus pusillus Dwarf Coastweed PH Asteraceae Ambrosia chamissonis Beach-bur S Asteraceae Ambrosia psilostachya Western Ragweed PH Asteraceae Amsinckia spectabilis var. spectabilis Seaside Fiddleneck AH Boraginaceae Anagallis arvensis * Scarlet Pimpernel AH Primulaceae Anemopsis californica Yerba Mansa PH Saururaceae Apium graveolens * Wild Celery PH Apiaceae Artemisia biennis Biennial Wormwood BH Asteraceae Artemisia californica California Sagebrush S Asteraceae Artemisia douglasiana Douglas' Sagewort PH Asteraceae Artemisia dracunculus Wormwood PH Asteraceae Artemisia tridentata ssp. tridentata Big Sagebrush S Asteraceae Arundo donax * Giant Reed PG Poaceae Aster subulatus var. ligulatus Annual Water Aster AH Asteraceae Astragalus pycnostachyus ssp. lanosissimus Ventura Marsh Milkvetch PH Fabaceae Atriplex californica California Saltbush PH Chenopodiaceae Atriplex lentiformis ssp. breweri Big Saltbush S Chenopodiaceae Atriplex patula ssp. hastata Arrowleaf Saltbush AH Chenopodiaceae Atriplex patula Spear Saltbush AH Chenopodiaceae Atriplex semibaccata Australian Saltbush PH Chenopodiaceae Atriplex triangularis Spearscale AH Chenopodiaceae Avena barbata * Slender Oat AG Poaceae Avena fatua * Wild -
![Dieteria [Machaeranthera] Cansecens](https://docslib.b-cdn.net/cover/6633/dieteria-machaeranthera-cansecens-446633.webp)
Dieteria [Machaeranthera] Cansecens
HOARY TANSYASTER Dieteria (Machaeranthera) canescens (Pursh) Nutt. Asteraceae – Aster family Corey L. Gucker & Nancy L. Shaw | 2018 ORGANIZATION NOMENCLATURE Dieteria canescens (Pursh) Nutt. until recently Names, subtaxa, chromosome number(s), hybridization. (2010) was known as Machaeranthera canescens (Pursh) A. Gray (ITIS 2017; USDA NRCS 2017). This species belongs to the Astereae tribe of the Asteraceae family (Morgan Range, habitat, plant associations, elevation, soils. 2006) and will hereafter be referred to by its common name, hoary tansyaster. NRCS Plant Code. MACA2 (USDA NRCS 2017). Life form, morphology, distinguishing characteristics, reproduction. Subtaxa. The Flora of North America (Morgan 2006) recognizes ten varieties of hoary tansyaster: Dieteria canescens var. Growth rate, successional status, disturbance ecology, importance to canescens, ambigua, aristata, glabra, incana, animals/people. leucanthemifolia, nebraskana, sessiliflora, shastensis, and ziegleri. Current or potential uses in restoration. Synonyms (Morgan 2006). Dieteria canescens: Machaeranthera canescens (Pursh) A. Gray; Aster canescens Seed sourcing, wildland seed collection, seed cleaning, storage, Pursh testing and marketing standards. D. c. var. ambigua: M. canescens (Pursh) A. Gray var. ambigua B.L. Turner D. c. var. aristatus: A. canescens Pursh var. Recommendations/guidelines for producing seed. aristatus Eastwood; M. canescens (Pursh) A. Gray var. aristata (Eastwood) B.L. Turner; M. rigida Greene D. c. var. canescens: M. divaricata (Nuttall) Recommendations/guidelines for producing planting stock. Greene; M. laetevirens Greene; M. latifolia A. Nelson; M. pulverulenta (Nuttall) Greene; M. viscosa (Nuttall) Greene D. c. var. glabra: M. canescens (Pursh) A. Gray Recommendations/guidelines, wildland restoration successes/ var. glabra A. Gray; A. canescens Pursh var. failures. viridis A. Gray; M. linearis Greene D. -

Anr-May-2017
May/June 2017 WARD COUNTY AG REPORT A bi-monthly publication of the Ward County Extension Office AGENT’S NOTE DROUGHT OUTLOOK Well folks, I’m sad to say this will be the last newsletter you receive from me as Ag Agent. I’ve accepted an offer for a position in the midstream industry, and I’ve decided to accept. My last day on duty will be June 9th. I want to thank you all for your support and cooperation over the past four years. I’ll still be running cows north of town so I’ll still see ya’ll around. Regards, - Courtesy of the National Drought Mitigation Center LIVESTOCK PRODUCERS SHOULD BE AWARE OF SMALL-HEADED SNEEZEWEED BY: Kay Ledbetter 1 1 1 Caleb L. Eaton County Extension Agent – Ag/NR Ward County 3600 S. Stockton Ave, Suite J Courthouse Annex Monahans, Texas 79756 Phone: (432) 943-2682 Email: [email protected] One plant currently flowering across different parts of the state is Blog: http://wardcountyextension.blogspot.com/ poisonous and should be of concern to ranchers, according to a Texas A&M AgriLife Extension Service expert. DEMONSTRATION CORNER Small -headed sneezeweed, which falls in the sunflower family, is a native, warm-season annual that grows statewide except for the East Texas Piney Woods and extends into northern Mexico, said Dr. Barron Rector, AgriLife Extension range specialist in College Station. “Be aware that small-headed sneezeweed is very poisonous in the flowering stage to mainly sheep, but cattle, goats, mules and RANGE MONITORING ON COX RANCH horses are also susceptible,” Rector said. -
1 C M 2 B H 3 B M 4 C M 5 B L 6 B M 7 B M 8
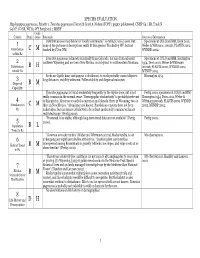
SPECIES EVALUATION Haplopappus pygmaeus, Priority 1. Tonestus pygmaeus (Torrey & Gray) A. Nelson (TOPY). pygmy goldenweed. CNHP G4 / SR, Track N G4 N?. CO SR, WY S1. WY Peripheral 2 MBNF Confi- Criteria Rank dence Rationale Sources of Information Distribution (see map below) is “nearly continuous,” so rating C was chosen, but Specimens at COLO and RM, Dorn 2001, 1 none of the pictures or descriptions really fit this species. Tracked by WY, but not Weber & Wittmann 2001ab, PLANTS 2002, Distribution C M tracked by CO or NM. WYNDD 2002. within R2 Tonestus pygmaeus is known principally from Colorado, but also from adjacent Specimens at COLO and RM, Harrington 2 southern Wyoming and northern New Mexico, and disjunct in southwestern Montana. 1954, Dorn 2001, Weber & Wittmann Distribution B H 2001ab, PLANTS 2002, WYNDD 2002, outside R2 MTNHP 2002. Seeds are lightly hairy and pappus is deciduous, so seeds probably cannot disperse Harrington 1954. 3 long distances; viability unknown. Pollen viability and dispersal unknown. Dispersal B M Capability Tonestus pygmaeus is found moderately frequently in the Alpine zone, but is not Fertig 2000, specimens at COLO and RM, really common in the normal sense. “Demographic stochasticity” is probably irrelevant Harrington 1954, Dorn 2001, Weber & 4 to this species. About 50 recorded occurrences in Colorado, three in Wyoming, two or Wittmann 2001ab, PLANTS 2002, WYNDD Abundance in C M three in New Mexico. “Abundance not known. Population censuses have not been 2002, MTNHP 2002. R2 undertaken, but the species is believed to be at least moderately common within its restricted range” (Fertig 2000). -

Reclassification of North American Haplopappus (Compositae: Astereae) Completed: Rayjacksonia Gen
AmericanJournal of Botany 83(3): 356-370. 1996. RECLASSIFICATION OF NORTH AMERICAN HAPLOPAPPUS (COMPOSITAE: ASTEREAE) COMPLETED: RAYJACKSONIA GEN. NOV.1 MEREDITH A. LANE2 AND RONALD L. HARTMAN R. L. McGregor Herbarium(University of Kansas NaturalHistory Museum Division of Botany) and Departmentof Botany,University of Kansas, Lawrence, Kansas 66047-3729; and Rocky MountainHerbarium, Department of Botany,University of Wyoming,Laramie, Wyoming82071-3165 Rayjacksonia R. L. Hartman& M. A. Lane, gen. nov. (Compositae: Astereae), is named to accommodate the "phyllo- cephalus complex," formerlyof Haplopappus Cass. sect. Blepharodon DC. The new combinationsare R. phyllocephalus (DC.) R. L. Hartman& M. A. Lane, R. annua (Rydb.) R. L. Hartman& M. A. Lane, and R. aurea (A. Gray) R. L. Hartman & M. A. Lane. This transfercompletes the reclassificationof the North American species of Haplopappus sensu Hall, leaving that genus exclusively South American.Rayjacksonia has a base chromosomenumber of x = 6. Furthermore,it shares abruptlyampliate disk corollas, deltatedisk style-branchappendages, and corolla epidermalcell type,among other features,with Grindelia, Isocoma, Olivaea, Prionopsis, Stephanodoria, and Xanthocephalum.Phylogenetic analyses of morphologicaland chloroplastDNA restrictionsite data, taken together,demonstrate that these genera are closely related but distinct. Key words: Astereae; Asteraceae; Compositae; Haplopappus; Rayjacksonia. During the past seven decades, taxonomic application lopappus sensu Hall (1928) are reclassifiedand are cur-