Reclassification of North American Haplopappus (Compositae: Astereae) Completed: Rayjacksonia Gen
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Checklist of the Vascular Flora of the Mary K. Oxley Nature Center, Tulsa County, Oklahoma
Oklahoma Native Plant Record 29 Volume 13, December 2013 A CHECKLIST OF THE VASCULAR FLORA OF THE MARY K. OXLEY NATURE CENTER, TULSA COUNTY, OKLAHOMA Amy K. Buthod Oklahoma Biological Survey Oklahoma Natural Heritage Inventory Robert Bebb Herbarium University of Oklahoma Norman, OK 73019-0575 (405) 325-4034 Email: [email protected] Keywords: flora, exotics, inventory ABSTRACT This paper reports the results of an inventory of the vascular flora of the Mary K. Oxley Nature Center in Tulsa, Oklahoma. A total of 342 taxa from 75 families and 237 genera were collected from four main vegetation types. The families Asteraceae and Poaceae were the largest, with 49 and 42 taxa, respectively. Fifty-eight exotic taxa were found, representing 17% of the total flora. Twelve taxa tracked by the Oklahoma Natural Heritage Inventory were present. INTRODUCTION clayey sediment (USDA Soil Conservation Service 1977). Climate is Subtropical The objective of this study was to Humid, and summers are humid and warm inventory the vascular plants of the Mary K. with a mean July temperature of 27.5° C Oxley Nature Center (ONC) and to prepare (81.5° F). Winters are mild and short with a a list and voucher specimens for Oxley mean January temperature of 1.5° C personnel to use in education and outreach. (34.7° F) (Trewartha 1968). Mean annual Located within the 1,165.0 ha (2878 ac) precipitation is 106.5 cm (41.929 in), with Mohawk Park in northwestern Tulsa most occurring in the spring and fall County (ONC headquarters located at (Oklahoma Climatological Survey 2013). -

Artemisia Californica Less
I. SPECIES Artemisia californica Less. [Updated 2017] NRCS CODE: Subtribe: Artemisiinae ARCA11 Tribe: Anthemideae (FEIS CODE: Family: Asteraceae ARCAL) Order: Asterales Subclass: Asteridae Class: Magnoliopsida flowering heads spring growth seedling, March 2009 juvenile plant photos A. Montalvo flowering plant, November 2005 mature plant with flower buds August 2010 A. Subspecific taxa None. Artemisia californica Less. var. insularis (Rydb.) Munz is now recognized as Artemisia nesiotica P.H. Raven (Jepson eFlora 2017). B. Synonyms Artemisia abrotanoides Nuttall; A. fischeriana Besser; A. foliosa Nuttall; Crossostephium californicum (Lessing) Rydberg (FNA 2017). C. Common name California sagebrush. The common name refers to its strong, sage-like aroma and endemism to California and Baja California. Other names include: coastal sage, coast sage, coast sagebrush (Painter 2016). D. Taxonomic relationships The FNA (2017) places this species in subgenus Artemisia . The molecular phylogeny of the genus has improved the understanding of relationships among the many species of Artemisia and has, at times, placed the species in subgenus Tridentadae; morphology of the inflorescences and flowers alone does not place this species with its closest relatives (Watson et al. 2002). The detailed phylogeny is not completely resolved (Hayat et al. 2009). E. Related taxa in region There are 18 species and a total of 31 taxa (including infrataxa) of Artemisia in southern California, all of which differ clearly from A. californica in habitat affinity, structure, or both (Munz 1974, Jepson eFlora 2017). Within subgenus Artemisia (as per FNA 2017), A. nesiotica from the Channel Islands is the most similar and was once considered part of A. californica ; it can be distinguished by its wider leaves with flat leaf margins (not rolled under). -

Field Guide for Managing Rabbitbrush in the Southwest
United States Department of Agriculture Field Guide for Managing Rabbitbrush in the Southwest Forest Southwestern Service Region TP-R3-16-31 February 2015 Cover Photos Top left: Green rabbitbrush, USDA Forest Service Top right: Green rabbitbrush flowers, Mary Ellen Harte, Bugwood.org Bottom left: Rubber rabbitbrush flowers, USDA Forest Service Bottom right: Rubber rabbitbrush, USDA Forest Service The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TTY). To file a complaint of discrimination, write to USDA, Director, Office of Civil Rights, 1400 Independence Avenue, SW, Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TTY). USDA is an equal opportunity provider and employer. Printed on recycled paper Green Rabbitbrush (Chrysothamnus viscidiflorus) Rubber Rabbitbrush (Chrysothamnus nauseosus, syn. Ericameria nauseosa) Sunflower family (Asteraceae) Green and rubber rabbitbrush are native shrubs that grow Some species are widespread geographically, and some are widely across western U.S. rangelands. Though they can restricted to a limited area. The specific rabbitbrush species appear as a weedy monoculture (especially following of concern should always be known before proceeding with disturbance), they are early colonizers and their presence management. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Notes on Variation and Geography in Rayjacksonia Phyllocephala (Asteraceae: Astereae)
Nesom, G.L., D.J. Rosen, and S.K. Lawrence. 2013. Notes on variation and geography in Rayjacksonia phyllocephala (Asteraceae: Astereae). Phytoneuron 2013-53: 1–15. Published 12 August 2013. ISSN 2153 733X NOTES ON VARIATION AND GEOGRAPHY IN RAYJACKSONIA PHYLLOCEPHALA (ASTERACEAE: ASTEREAE) GUY L. NESOM 2925 Hartwood Drive Fort Worth, Texas 76109 [email protected] DAVID J. ROSEN Department of Biology Lee College Baytown, Texas 77522-0818 [email protected] SHIRON K. LAWRENCE Department of Biology Lee College Baytown, Texas 77522-0818 ABSTRACT Inflorescences of Rayjacksonia phyllocephala in the disjunct Florida population system are characterized by heads on peduncles with leaves mostly reduced to linear bracts; heads in inflorescences of the Mexico-Texas-Louisiana system are immediately subtended by relatively unreduced leaves. The difference is consistent and justifies recognition of the Florida system as R. phyllocephala var. megacephala (Nash) D.B. Ward. Scattered waifs between the two systems are identified here as one or the other variety, directly implying their area of origin. In the eastern range of var. phyllocephala , at least from Brazoria County, Texas, eastward about 300 miles to central Louisiana, leaf margins vary from entire to deeply toothed-spinulose. In contrast, margins are invariably toothed-spinulose in var. megacephala as well as in the rest of southeastern Texas (from Brazoria County southwest) into Tamaulipas, Mexico. In some of the populations with variable leaf margins, 70-95% of the individuals have entire to mostly entire margins. KEY WORDS : Rayjacksonia , morphological variation, leaf margins, disjunction, waifs Rayjacksonia phyllocephala (DC.) Hartman & Lane (Gulf Coast camphor-daisy) is an abundant and conspicuous species of the shore vegetation around the Gulf of Mexico. -

Literature Cited
Literature Cited Robert W. Kiger, Editor This is a consolidated list of all works cited in volumes 19, 20, and 21, whether as selected references, in text, or in nomenclatural contexts. In citations of articles, both here and in the taxonomic treatments, and also in nomenclatural citations, the titles of serials are rendered in the forms recommended in G. D. R. Bridson and E. R. Smith (1991). When those forms are abbre- viated, as most are, cross references to the corresponding full serial titles are interpolated here alphabetically by abbreviated form. In nomenclatural citations (only), book titles are rendered in the abbreviated forms recommended in F. A. Stafleu and R. S. Cowan (1976–1988) and F. A. Stafleu and E. A. Mennega (1992+). Here, those abbreviated forms are indicated parenthetically following the full citations of the corresponding works, and cross references to the full citations are interpolated in the list alphabetically by abbreviated form. Two or more works published in the same year by the same author or group of coauthors will be distinguished uniquely and consistently throughout all volumes of Flora of North America by lower-case letters (b, c, d, ...) suffixed to the date for the second and subsequent works in the set. The suffixes are assigned in order of editorial encounter and do not reflect chronological sequence of publication. The first work by any particular author or group from any given year carries the implicit date suffix “a”; thus, the sequence of explicit suffixes begins with “b”. Works missing from any suffixed sequence here are ones cited elsewhere in the Flora that are not pertinent in these volumes. -

Phytologia an International Journal to Expedite
PHYTOLOGIA An intern ational jou rnal to ex edite la n t s stematic b to eo ra bi l p p y , p y g g p ca and ecological pu blication 1 S e ember 1 1 Vo l. 7 pt 99 CONTENTS TRE M N w ll n n n t r c l fl r . A ne CUA CASAS . c o o o o o o , J , is e a e us tes e pi a a XX species of Humiriastru m 1 65 R T F w f S . rm l rr t n f t c th in the th rn OSS . o co c o o o c t o , , a e i spe i i epi e s s u e . N N H N A eratina nd w f B artlettina ROB S O . o t on a ne c o / I , , es g a spe ies (Eupato rieae : Asteraceae) 1 7 1 /R )BIN N H N w w f C SO . e c and ne comb n on o Crit oniinae from , , spe ies i ati s Meso ame ri ca (Eupato rieae : Asteraceae) 1 76 ’ R B N N H Tw w f Fleiscbman m a fr m M r O S O . o ne c o o m c / I , , spe ies o es a e i a (Eupatorieae : As t eraceae) 1 8 1 /ROBIN N Tw w M le a M SO H. o ne c o f i an i in m r c m , , spe ies esoa e i a (Eupatorieae : As te raceae) 1 84 E AL N D L f f a a r i S C O A F . -

Pdf Clickbook Booklet
183 Liliaceae Yucca brevifolia joshua tree 5 184 Liliaceae Yucca schidigera Mohave yucca 99 1 Flora of New Dixie Mine Road Area, west of Landers Achnatherum 185 Poaceae ~ desert needlegrass 20 2 # Plants speciosum # JM Family ID? Scientific Name (*)Common Name Bloom #Vch six-weeks Obs'd 186 Poaceae Aristida adscensionis 50 99 22Oct10 three-awn 1 Pteridaceae Cheilanthes covillei beady lipfern 1 Aristida purpurea var. 187 Poaceae ssp Nealley three-awn 1 1 4 nealleyi Pentagramma 2 Pteridaceae ~ triangularis ssp. goldback fern 1 Bouteloua barbata var. 188 Poaceae six-weeks grama 20 50 10 triangularis barbata 3 Cupressaceae Juniperus californica California juniper 1 Bromus madritensis ssp. 189 Poaceae *red brome 99 2 rubens 4 Ephedraceae Ephedra californica desert tea 30 1 190 Poaceae Bromus trinii *Chilean chess 3 5 Ephedraceae Ephedra nevadensis Nevada ephedra 3 191 Poaceae Distichlis spicata saltgrass 1 6 Ephedraceae Ephedra viridis green ephedra 1 Elymus elymoides ssp. 7 Pinaceae Pinus monophylla pinyon pine 2 192 Poaceae squirreltail 2 elymoides 8 Amaranthaceae Amaranthus fimbriatus fringed amaranth 99 99 11 193 Poaceae Erioneuron pulchellum fluff grass 25 9 Apiaceae Lomatium mohavense Mojave lomatium 1 Hordeum vulgare var. 194 Poaceae *cultivated barley 1 Acamptopappus trifurcatum 10 Asteraceae ~ sphaerocephalus var. goldenhead 5 3 195 Poaceae Melica frutescens tall melica 2 sphaerocephalus 196 Poaceae Melica imperfecta coast-range melic 3 11 Asteraceae Adenophyllum cooperi Cooper's dogweed 5 20 2 197 Poaceae Muhlenbergia rigens deergrass 2 12 Asteraceae Ambrosia acanthicarpa bur-ragweed 1 198 Poaceae Pleuraphis rigida big galleta 10 99 11 13 Asteraceae Ambrosia dumosa burroweed 5 99 12 Poa secunda ssp. -
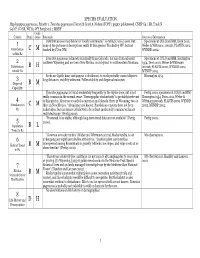
1 C M 2 B H 3 B M 4 C M 5 B L 6 B M 7 B M 8
SPECIES EVALUATION Haplopappus pygmaeus, Priority 1. Tonestus pygmaeus (Torrey & Gray) A. Nelson (TOPY). pygmy goldenweed. CNHP G4 / SR, Track N G4 N?. CO SR, WY S1. WY Peripheral 2 MBNF Confi- Criteria Rank dence Rationale Sources of Information Distribution (see map below) is “nearly continuous,” so rating C was chosen, but Specimens at COLO and RM, Dorn 2001, 1 none of the pictures or descriptions really fit this species. Tracked by WY, but not Weber & Wittmann 2001ab, PLANTS 2002, Distribution C M tracked by CO or NM. WYNDD 2002. within R2 Tonestus pygmaeus is known principally from Colorado, but also from adjacent Specimens at COLO and RM, Harrington 2 southern Wyoming and northern New Mexico, and disjunct in southwestern Montana. 1954, Dorn 2001, Weber & Wittmann Distribution B H 2001ab, PLANTS 2002, WYNDD 2002, outside R2 MTNHP 2002. Seeds are lightly hairy and pappus is deciduous, so seeds probably cannot disperse Harrington 1954. 3 long distances; viability unknown. Pollen viability and dispersal unknown. Dispersal B M Capability Tonestus pygmaeus is found moderately frequently in the Alpine zone, but is not Fertig 2000, specimens at COLO and RM, really common in the normal sense. “Demographic stochasticity” is probably irrelevant Harrington 1954, Dorn 2001, Weber & 4 to this species. About 50 recorded occurrences in Colorado, three in Wyoming, two or Wittmann 2001ab, PLANTS 2002, WYNDD Abundance in C M three in New Mexico. “Abundance not known. Population censuses have not been 2002, MTNHP 2002. R2 undertaken, but the species is believed to be at least moderately common within its restricted range” (Fertig 2000). -

The Sabal May 2017
The Sabal May 2017 Volume 34, number 5 In this issue: Native Plant Project (NPP) Board of Directors May program p1 below Texas at the Edge of the Subtropics— President: Ken King by Bill Carr — p 2-6 Vice Pres: Joe Lee Rubio Native Plant Tour Sat. May 20 in Harlingen — p 7 Secretary: Kathy Sheldon Treasurer: Bert Wessling LRGV Native Plant Sources & Landscapers, Drew Bennie NPP Sponsors, Upcoming Meetings p 7 Ginger Byram Membership Application (cover) p8 Raziel Flores Plant species page #s in the Sabal refer to: Carol Goolsby “Plants of Deep South Texas” (PDST). Sande Martin Jann Miller Eleanor Mosimann Christopher Muñoz Editor: Editorial Advisory Board: Rachel Nagy Christina Mild Mike Heep, Jan Dauphin Ben Nibert <[email protected]> Ken King, Betty Perez Ann Treece Vacek Submissions of relevant Eleanor Mosimann NPP Advisory Board articles and/or photos Dr. Alfred Richardson Mike Heep are welcomed. Ann Vacek Benito Trevino NPP meeting topic/speaker: "Round Table Plant Discussion" —by NPP members and guests Tues., April 23rd, at 7:30pm The Native Plant Project will have a Round Table Plant Discussion in lieu of the usual PowerPoint presentation. We’re encouraging everyone to bring a native plant, either a cutting or in a pot, to be identified and discussed at the meeting. It can be a plant you are unfamiliar with or something that you find remarkable, i.e. blooms for long periods of time or has fruit all winter or is simply gor- geous. We will take one plant at a time and discuss it with the entire group, inviting all comments about your experience with that native. -

References and Appendices
References Ainley, D.G., S.G. Allen, and L.B. Spear. 1995. Off- Arnold, R.A. 1983. Ecological studies on six endan- shore occurrence patterns of marbled murrelets gered butterflies (Lepidoptera: Lycaenidae): in central California. In: C.J. Ralph, G.L. Hunt island biogeography, patch dynamics, and the Jr., M.G. Raphael, and J.F. Piatt, technical edi- design of habitat preserves. University of Cali- tors. Ecology and Conservation of the Marbled fornia Publications in Entomology 99: 1–161. Murrelet. USDA Forest Service, General Techni- Atwood, J.L. 1993. California gnatcatchers and coastal cal Report PSW-152; 361–369. sage scrub: the biological basis for endangered Allen, C.R., R.S. Lutz, S. Demairais. 1995. Red im- species listing. In: J.E. Keeley, editor. Interface ported fire ant impacts on Northern Bobwhite between ecology and land development in Cali- populations. Ecological Applications 5: 632-638. fornia. Southern California Academy of Sciences, Allen, E.B., P.E. Padgett, A. Bytnerowicz, and R.A. Los Angeles; 149–169. Minnich. 1999. Nitrogen deposition effects on Atwood, J.L., P. Bloom, D. Murphy, R. Fisher, T. Scott, coastal sage vegetation of southern California. In T. Smith, R. Wills, P. Zedler. 1996. Principles of A. Bytnerowicz, M.J. Arbaugh, and S. Schilling, reserve design and species conservation for the tech. coords. Proceedings of the international sym- southern Orange County NCCP (Draft of Oc- posium on air pollution and climate change effects tober 21, 1996). Unpublished manuscript. on forest ecosystems, February 5–9, 1996, River- Austin, M. 1903. The Land of Little Rain. University side, CA. -

Grindeliae Herba Grindelia 2015
ONLINE SERIES MONOGRAPHS The Scientific Foundation for Herbal Medicinal Products Grindeliae herba Grindelia 2015 www.escop.com The Scientific Foundation for Herbal Medicinal Products GRINDELIAE HERBA Grindelia 2015 ESCOP Monographs were first published in loose-leaf form progressively from 1996 to 1999 as Fascicules 1-6, each of 10 monographs © ESCOP 1996, 1997, 1999 Second Edition, completely revised and expanded © ESCOP 2003 Second Edition, Supplement 2009 © ESCOP 2009 ONLINE SERIES ISBN 978-1-901964-34-9 Grindeliae herba - Grindelia © ESCOP 2015 Published by the European Scientific Cooperative on Phytotherapy (ESCOP) Notaries House, Chapel Street, Exeter EX1 1EZ, United Kingdom www.escop.com All rights reserved Except for the purposes of private study, research, criticism or review no part of this text may be reproduced, stored in a retrieval system or transmitted, in any form or by any means, without the written permission of the publisher. Important Note: Medical knowledge is ever-changing. As new research and clinical experience broaden our knowledge, changes in treatment may be required. In their efforts to provide information on the efficacy and safety of herbal drugs and herbal preparations, presented as a substantial overview together with summaries of relevant data, the authors of the material herein have consulted comprehensive sources believed to be reliable. However, in view of the possibility of human error by the authors or publisher of the work herein, or changes in medical knowledge, neither the authors nor the publisher, nor any other party involved in the preparation of this work, warrants that the information contained herein is in every respect accurate or complete, and they are not responsible for any errors or omissions or for results obtained by the use of such information.