Sheepmeat Flavour and Odour: a Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Where's the Beef? Communicating Vegetarianism in Mainstream America
Where’s the Beef? Communicating Vegetarianism in Mainstream America ALLISON WALTER Produced in Mary Tripp’s Spring 2013 ENC 1102 Introduction “Engaging in non-mainstream behavior can be challenging to negotiate communicatively, especially when it involves the simple but necessary task of eating, a lifelong activity that is often done in others’ company,” argue researchers Romo and Donovan-Kicken (405). This can be especially true for vegetarians in America. The American view of a good and balanced meal includes a wide array of meats that have become standard on most all of the country's dinner tables. When it comes to eating in American society today, fruits and vegetables come to mind as side dishes or snacks, not the main course. Vegetarians challenge this expectation of having meat as an essential component of survival by adopting a lifestyle that no longer conforms to the norms of society as a whole. Although vegetarians can be seen as “healthy deviants”—people who violate social norms in relatively healthy ways—they are faced with the burden of stigmatization by those who cannot see past their views of conformity (Romo and Donovan-Kicken 405). This can lead to a continuous battle for vegetarians where they are questioned and scrutinized for their decisions only because they are not the same as many of their peers. The heaviest load that individuals who decide to deviate from the norms of a given society have to carry is attempting to stay true to themselves in a world that forces them to fit in. Culture, Society, and Food One of the largest and most complex factors that contribute to food choice is society and the cultures within that society that certain groups of people hold close to them (Jabs, Sobal, and Devine 376). -

Greater China Beef & Sheepmeat Market Snapshot
MARKET SNAPSHOT l BEEF & SHEEPMEAT Despite being the most populous country in the world, the proportion of Chinese consumers who can regularly afford to buy Greater China high quality imported meat is relatively small in comparison to more developed markets such as the US and Japan. However, (China, Hong Kong and Taiwan) continued strong import demand for premium red meat will be driven by a significant increase in the number of wealthy households. Focusing on targeted opportunities with a differentiated product will help to build preference in what is a large, complex and competitive market. Taiwan and Hong Kong are smaller by comparison but still important markets for Australian red meat, underpinned by a high proportion of affluent households. China meat consumption 1 2 Population Household number by disposable income per capita3 US$35,000+ US$75,000+ 1,444 13% million 94.3 23% 8% 50 kg 56% 44.8 China grocery spend4 10.6 24.2 3.3 5.9 million million 333 26 3.0 51 16.1 Australia A$ Australia Korea China Korea Australia China US US China US Korea 1,573 per person 1,455 million by 2024 5% of total households 0.7% of total households (+1% from 2021) (9% by 2024) (1% by 2024) Australian beef exports to China have grown rapidly, increasing 70-fold over the past 10 years, with the country becoming Australia’s largest market in 2019. Australian beef Australian beef Australia’s share of Australian beef/veal exports – volume5 exports – value6 direct beef imports7 offal cuts exports8 4% 3% 6% 6% 3% Tripe 21% 16% Chilled grass Heart 9% A$ Chilled grain Chilled Australia Tendon Other Frozen grass Frozen 10% 65 71% Tail 17% countries million Frozen grain 84% Kidney 67% Other Total 303,283 tonnes swt Total A$2.83 billion Total 29,687 tonnes swt China has rapidly become Australia’s single largest export destination for both lamb and mutton. -

Information on How to Buy Lamb
LAMB Lamb is produced from animals less than a year old. Since the quality of lamb varies according to the age of the animal, it is advisable to buy lamb that has been USDA-graded. USDA Prime: Prime grade lamb is very high in tenderness, juiciness, and flavor. It has moderate marbling, which enhances both flavor and juiciness. Prime chops and roasts are excellent for dry-heat cooking (broiling and roasting). USDA Choice: Choice grade lamb has slightly less marbling than Prime, but still is of very high quality. Choice chops and roasts also are very tender, juicy, and flavorful and suited to dry-heat cooking. Lower grades of lamb and mutton (USDA Good, Utility, and Cull) are seldom marked with the grade if sold at retail. Most cuts of USDA Prime and Choice lamb - including shoulder cuts - are tender and can be oven roasted, broiled, or pan broiled. A leg of lamb graded Choice or Prime, for example, is delectable when oven roasted. The less tender cuts - the breast, riblets, neck, and shank - can be braised slowly to make excellent (and tender) lamb dishes. Meat from older sheep is called yearling mutton or mutton and, if it is graded, these words will be stamped on the meat along with the shield-shaped grade mark. Grades for yearling mutton and mutton are the same as for lamb, except that mutton does not qualify for the Prime grade and the Cull grade applies only to mutton. The best way to identify lamb cuts is with the lamb carcass chart shown above. -

United States Standards for Grades of Lamb, Yearling Mutton, and Mutton Carcasses
United States Department of Agriculture United States Standards Agricultural Marketing for Grades of Service Livestock Lamb, Yearling Mutton, and and Seed Division Mutton Carcasses Effective date July 6, 1992 United States Standards for Grades of Lamb, Yearling Mutton, and Mutton Carcasses The following is a reprint of the Official United States Standards for the Grades of Lamb, Yearling Mutton, and Mutton Carcasses promulgated by the Secretary of Agriculture under the Agricultural Marketing Act of 1946 (60 Stat. 1087; 7 U.S.C. 1621-1627) as amended and related authority in the annual appropriation acts for the Department of Agriculture. The standards are reprinted with amendments effective July 6, 1992. Development of the Standards The official standards for grades of lamb and mutton carcasses were initially promulgated and made effective on February 16, 1931. The standards were amended in October 1940 (Amendment No. 1 to S.R.A. 123) so as to change the grade designations Medium and common to Commercial and Utility, respectively. In April 1951, the official standards were again amended (Amendment No. 2 to S.R.A. 123). By this amendment, Prime and Choice grades were combined and designated as Prime. The Good grade was renamed Choice, which also became the highest grade for carcasses of mutton older than yearlings. The top two-thirds of the Commercial grade was combined with the top two-third of the Utility grade and designated as Utility, thereby eliminating the Commercial grade name. The lower one-third of the Utility grade was combined with the Cull grade and designated as Cull. This amendment also provided for reflecting the minimum requirements for each grade, specified the grade requirements for varying degrees of maturity, and clarified the method for differentiating between lamb, yearling mutton, and mutton carcasses. -

Spicy Beef Tongue Stew
Spicy Beef Tongue Stew Serves six 1 beef tongue – trimmed 1 celery stalk 1 small carrot – halved 2 garlic cloves – peeled and smashed + 1 for the chilies 4 fresh thyme sprigs 1 small white onion – peeled and halved + ½ for the chilies 1 chili pepper – halved 6 red chilies (aji Colorado or aji Panka) or 3 Guajillos, 3 Ancho and 3 chile Arbol 1 TBS cumin 1 tsp. salt 1/8 cup canola oil ¼ cup peas (can be thawed frozen ones or fresh) 1 large carrot – peeled and cut into sticks Salt & Pepper Rinse the trimmed beef tongue and place it in a pressure cooker. Add the celery, onion, carrot, garlic cloves, thyme and chili pepper. Cook for about 1 hour – after the pressure cooker starts making noise. While the tongue is cooking – stem the dry chili pods. Cut them in the middle and seed them. Char them by placing them directly on an open flame. If you do not have a gas stove, go ahead and press them down on a dry, hot skillet until they blister. Place the charred chilies in a bowl and cover with water. Soak for about 25 to 30 minutes. Once soaked, place them in a blender with the remaining onion, garlic and cumin. Add about 1 cup of the soaking liquid and blend until you have a smooth paste with no chili chunks. Once the tongue is cooked – cool down the pressure cooker completely and remove them. Do not discard the cooking liquid. Cool them down and peel them by pulling on the skin and membrane on the meat. -

12 Steak Tartare
November 22nd, 2018 3 Course Menu, $42 per person ( tax & gratuity additional ) ( no substitutions, no teal deals, thanksgiving day menu is a promotional menu ) Salad or Soup (choice, descriptions below) Caesar Salad, Wedge Salad, Lobster Bisque or Italian White Bean & Kale Entrée (choice, descriptions below) Herb Roasted Turkey, Local Flounder, Faroe Island Salmon or Braised Beef Short Rib Dessert (choice) Dark Chocolate Mousse Cake, Old Fashioned Carrot Cake, Pumpkin Cheesecake, Cinnamon Ice Cream, Vanilla Ice Cream or Raspberry Sorbet A LA CARTE MENU APPETIZERS ENTREES Butternut Squash & Gouda Arancini … 13 Herb Roasted Fresh Turkey … 32 (5) risotto arancini, asparagus pesto, truffle oil, mashed potato, herbed chestnut stuffing, fried sage, aged pecorino Romano glazed heirloom carrots, haricot vert, truffled brown gravy, cranberry relish Smoked Salmon Bruschetta … 12 house smoked Faroe Island salmon, lemon aioli, Local Flounder … 32 micro salad with shallot - dill vinaigrette, crispy capers warm orzo, sweet corn, sun dried tomato, edamame & vidalia, sautéed kale, bell pepper-saffron jam, basil pesto Steak Tartare … 14 beef tenderloin, sous vide egg yolk, caper, Faroe Island Salmon* … 32 shallot, lemon emulsion, parmesan dust, garlic loaf toast roasted spaghetti squash, haricot vert & slivered almonds, carrot puree, garlic tomato confit, shallot dill beurre blanc Grilled Spanish Octopus … 14 gigante bean & arugula sauté, grape tomatoes, Boneless Beef Short Rib … 32 salsa verde, aged balsamic reduction oyster mushroom risotto, sautéed -

Sensory Evaluation of Various Lamb Meat Foods by Judging Consumers
KUKOVICS & NÉMETH: SENSORY EVALUATION OF LAMB MEAT FOODS BY CONSUMERS Acta Alimentaria, Vol. 43 (2), pp. 254–263 (2014) DOI: 10.1556/AAlim.43.2014.2.9 SENSORY EVALUATION OF VARIOUS LAMB MEAT FOODS BY JUDGING CONSUMERS S. KUKOVICS* and T. NÉMETH Research Institute for Animal Breeding and Nutrition, 2053 Herceghalom, Gesztenyés u. 1. Hungary (Received: 6 November 2012; accepted: 24 February 2013) Four kinds of lamb meat foods (Wiener sausage, ham, roasted meat, and roasted leg) were developed based on 8 different genotypes to improve lamb meat quality and quantity, and to study the differences among them. The stringiness, flavour, and odour of the products were judged in food sensory evaluation by a total of 265 (60% male and 40% female) randomly selected consumers. In stringiness and flavour five (from 1 to 5), and concerning odour three (from 1 to 3) categories were created according to decreasing quality. The consumers represented different age groups: below 20 years, between 21–30, 31–40, 41–50, 51–60, 61–70, and above 70 years. The groups of consumers were involved in sheep farming, other animal farming, other agriculture sector, industry, catering, education, other service; administrative department of the state, were students, and others. Analyses of variance were calculated to estimate the effects of the genotype of lamb, the gender, age, and occupation of judging consumers on the sensory evaluation of the meat foods. In conclusion, new lamb meat foods were highly appreciated by consumers, who made definite distinctions among genotypes according to flavour, stinginess, and odour. There were significant interactions between gender, age, and occupation of consumers as well as sensory evaluation of various lamb meat foods. -
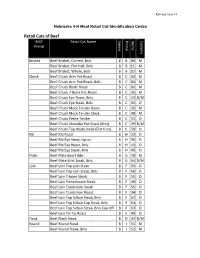
Retail Cuts of Beef BEEF Retail Cut Name Specie Primal Name Cookery Primal
Revised June 14 Nebraska 4-H Meat Retail Cut Identification Codes Retail Cuts of Beef BEEF Retail Cut Name Specie Primal Name Cookery Primal Brisket Beef Brisket, Corned, Bnls B B 89 M Beef Brisket, Flat Half, Bnls B B 15 M Beef Brisket, Whole, Bnls B B 10 M Chuck Beef Chuck Arm Pot-Roast B C 03 M Beef Chuck Arm Pot-Roast, Bnls B C 04 M Beef Chuck Blade Roast B C 06 M Beef Chuck 7-Bone Pot-Roast B C 26 M Beef Chuck Eye Roast, Bnls B C 13 D/M Beef Chuck Eye Steak, Bnls B C 45 D Beef Chuck Mock Tender Roast B C 20 M Beef Chuck Mock Tender Steak B C 48 M Beef Chuck Petite Tender B C 21 D Beef Chuck Shoulder Pot Roast (Bnls) B C 29 D/M Beef Chuck Top Blade Steak (Flat Iron) B C 58 D Rib Beef Rib Roast B H 22 D Beef Rib Eye Steak, Lip-on B H 50 D Beef Rib Eye Roast, Bnls B H 13 D Beef Rib Eye Steak, Bnls B H 45 D Plate Beef Plate Short Ribs B G 28 M Beef Plate Skirt Steak, Bnls B G 54 D/M Loin Beef Loin Top Loin Steak B F 59 D Beef Loin Top Loin Steak, Bnls B F 60 D Beef Loin T-bone Steak B F 55 D Beef Loin Porterhouse Steak B F 49 D Beef Loin Tenderloin Steak B F 56 D Beef Loin Tenderloin Roast B F 34 D Beef Loin Top Sirloin Steak, Bnls B F 62 D Beef Loin Top Sirloin Cap Steak, Bnls B F 64 D Beef Loin Top Sirloin Steak, Bnls Cap Off B F 63 D Beef Loin Tri-Tip Roast B F 40 D Flank Beef Flank Steak B D 47 D/M Round Beef Round Steak B I 51 M Beef Round Steak, Bnls B I 52 M BEEF Retail Cut Name Specie Primal Name Cookery Primal Beef Bottom Round Rump Roast B I 09 D/M Beef Round Top Round Steak B I 61 D Beef Round Top Round Roast B I 39 D Beef -

Sheep Skill-A-Thon Booklet
2016-2017 Citrus County Fair Sheep Skill-a-Thon Sheep Skill-a-thon 1 INTRODUCTION This manual has been developed by the Animal Sciences Department Faculty at the University of Florida as a study guide for the Sheep Skill-a-thon. The topic for this year’s Skill-a-thon is Products and Marketing. The Citrus County Fair recognizes that agricultural education instructors, parents, 4H agents and leaders provide the traditional and logical instructional link between youth, their livestock projects and current trends in the animal agriculture industry. PLEASE NOTE: This manual is provided as a study guide for the Skill-a-thon competition and should be used as an additional aid to ongoing educational programs. Sections are labeled Junior, Intermediate & Senior, Intermediate & Senior, or Senior to help exhibitors and educators identify which materials are required for each age level. The knowledge and skills vary by age group and may include: Juniors (age 8-10 as of September 1, 2016) By Products, Wholesale cuts & Primal Intermediates (age 11-13 as of September 1, 2016) all of the above plus... Retail Cuts Cookery Seniors (age 14 and over as of September 1, 2016) all of the above plus.... Sheep/Lamb Evaluation Wool Grades Quality Assurance Skeletal Anatomy GOOD LUCK! Sheep Skill -a-thon 2 Products and Marketing Youth livestock projects focus on the selection, raising, showing and often selling of animals. By virtue of their participation in livestock projects, youth become part of an industry that provides food and fiber for the world. Steps involved in the movement of animals and animal products from producer to consumer are known as processing and marketing. -

HOW MUCH MEAT to EXPECT from a BEEF CARCASS Rob Holland, Director Center for Profitable Agriculture
PB 1822 HOW MUCH MEAT TO EXPECT FROM A BEEF CARCASS Rob Holland, Director Center for Profitable Agriculture Dwight Loveday, Associate Professor Department of Food Science and Technology Kevin Ferguson UT Extension Area Specialist-Farm Management University of Tennessee Institute of Agriculture CONTENTS 2...Introduction 3...Dressing Percentage 5...Chilled Carcass and Primal Cuts 6...Sub-primal Meat Cuts 6...Factors Affecting Yield of Retail Cuts 7...Average Amount of Meat from Each Sub-primal Cut 9...Summary University of Tennessee Institute of Agriculture Introduction Consumers who buy a live animal from a local cattle producer for custom processing are often surprised. Some are surprised at the quantity of meat and amount of freezer space they need. Others may be surprised that they did not get the entire live weight of the animal in meat cuts. The amount of meat actually available from a beef animal is a frequent source of misunderstanding between consumers, processors and cattle producers. This document provides information to assist in the understanding of how much meat to expect from a beef carcass. The information provided here should be helpful to consumers who purchase a live animal for freezer beef and to cattle producers involved in direct and retail meat marketing. 2 University of Tennessee Institute of Agriculture How Much Meat to Expect from a Beef Carcass Dressing Percentage One of the terms used in the cattle and meat cutting industry that often leads to misunderstanding is dressing percentage. The dressing percentage is the portion of the live animal weight that results in the hot carcass. -

Beef Breakdown.Indd
Your Beef Breakdown, Explained If you have ever purchased a quarter, a side or a whole 14- to 21-day period. During this aging process the meat Loin and Round, all possess di erent beef carcass, chances are you have wondered why you have develops avor, and most importantly, becomes more taste characteristics because they are received less meat than expected. e average weight of a tender. Unfortunately, a small amount of weight is lost made up of di erent muscles with live steer or heifer ready for harvest is 1,300 pounds! So during the aging process due to water evaporation. di erent tenderness levels, di erent how much of this product should you expect to receive in Carcass to Cuts fat contents and varying avor pro les. edible meat products? For instance, the Chuck and Round After the carcass is properly aged, it is ready to be are most commonly seen in roast Steer to Carcass broken down into retail cuts. On average, 21 percent of form, but Round Roasts are much In order to change a 1,300-pound beef animal into edible each carcass is inedible bone, fat and connective tissue. leaner (have less fat) than those from meat product, butchers rst have to convert it into a Once the carcass is fabricated and inedible objects are the Chuck and therefore will have a carcass by removing the hide, head and internal organs. removed, a whole carcass will yield about 639 pounds of less intense avor. e Rib and Loin On average, only 62 percent of the animal’s original edible beef product. -

Tongue by Jennifer Mclagan from Odd Bits: How to Cook
Tongue By Jennifer McLagan from Odd Bits: How to Cook the Rest of the Animal How to Choose: The world of tongues is vast. Duck tongues, popular in Asian cuisines, are tiny and have a small bone that makes them fiddly. In this section, I limit myself to beef, veal, lamb, and pork tongues; if you have access to game animals, don’t hesitate to use their tongues in these recipes. The bigger the tongue, the coarser its texture, so I prefer tongues around 31/3 pounds /1.5 kg or less, as their texture is finer. While veal tongues are the most prized, beef, lamb, and pork tongues are all worth eating. Tongues can be bought fresh, frozen, brined, and sometimes smoked. Often your only choice will be a frozen one, especially with beef or veal tongue. The color of a tongue can vary from pink with a gray cast to almost all gray. Sometimes the skin of the tongue is quite mottled, often with odd dark spots. The color and the dark patches are no indication of quality; they are just a result of the animal’s breed. Tongues have a thick, bumpy skin, and often there is fat and gristle still attached at the base of the tongue, none of which is very appealing and all of which is easily removed after the tongue is poached. Besides availability, taste and size will influence your choice. Veal and lamb tongues are the mildest in taste, followed by beef and pork tongue. Pork tongue often comes with the head, and I prefer it in headcheese rather than by itself.