Dictyodora and Associated Trace Fossils from the Palaeozoic Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Burgfest Anlässlich 750 Jahre Bad Blankenburg Siehe Seite 11
1992 2017 25 Jahre Mitteilungsblatt der Greifenstein-Freunde Bad Blankenburg e.V. 25. Jahrgang Juni 2017 Ausgabe Nr. 46 Burgfest anlässlich 750 Jahre Bad Blankenburg siehe Seite 11 Greifenstein-Bote Inhalt Unsere Jubilare Für den Inhalt der Beiträge zeichnen derenVerfasser verantwortlich. Von Dieter Krause; Fotos: Greifenstein-Freunde Unsere Jubilare im II. Quartal 2017 ......................... Seite 2 Im 2. Quartal konnten folgende Greifenstein-Freunde besondere Jubiläen feiern: April - April .................................... Seite 2 750 Jahre Bad Blankenburg ����������� Seite 3 Durchführung der Walpurgisnacht . Seite 3 25 Jahre Greifenstein Bote. ............ Seite 4 Dank an unsere Spender ................. Seite 4 40 Jahre Bier- und Burgenstraße .... Seite 5 Ursprung des Spottnamens Teil 2 .. Seite 6 Arbeitseinsätze auf Burg Greifenstein ..................... Seite 8 Verabschiedung der Standesbeamtin ........................ Seite 9 Wieder eine Sorge weniger .......... Seite 10 Nachbetrachtung zum Burgfest .... Seite 11 Jungautoren zu Gast * Prof. Dr. Eberhard Weise am 18. Mai * Jürgen Michele am 1. Juni auf Burg Greifenstein ................... Seite 12 in Monheim den 90. Geburtstag, in Bad Blankenburg den 55. Geburtstag, Bildergalerie Himmelfahrt ........... Seite 12 Vereinsmitglied seit 20 Jahren, und Mitglied seit 2016, sowie Wir wünschen unseren Jubilaren al- les Gute und bedanken uns für ihre Impressum langjährige aktive Förderung bzw. Mitarbeit. Gesundheit und Wohlerge- Titelfoto: hen mögen dazu beitragen, dass noch Werner Nitschke viele Jahre eine konstruktive und gute Zusammenarbeit möglich ist, um das Redaktion: Wahrzeichen der Stadt Bad Blanken- Marcella Nitschke burg in einem solch sehenswürdigen Auf dem Sande 2 Zustand zu erhalten. 07422 Bad Blankenburg Wir erinnern: Am 10. Juni wäre Dieter Tel. 036741 2001 Klotz 80 Jahre alt geworden. Seine Mitarbeit E-Mail: [email protected] im Verein war im Besonderen in der Bilddo- * Marcella Nitschke am 7. -

Planispiral Burrows from a Recent Lacustrine Beach, Gander Lake, Newfoundland
Planispiral Burrows from a Recent Lacustrine Beach, Gander Lake, Newfoundland ROBERT B. M AC NAUGHTON Geological Survey of Canada, 3303 – 33 Street NW, Calgary, Alberta T2L 2A7 Canada; e-mail: [email protected] MacNaughton, Robert B. 2003. Planispiral burrows from a Recent lacustrine beach, Gander Lake Newfoundland. Canadian Field-Naturalist 117(4): 577-581. Simple horizontal, planispiral burrows occur in Recent beach sediments on the south shore of Gander Lake, Newfoundland. The burrows apparently represent deposit feeding, possibly by an arthropod. This occurrence extends the known environ- mental range of such traces into lacustrine settings and illustrates that such behaviour is not exclusive to marine settings. Key Words: ichnology, lacustrine, Recent, palaeoecology, Newfoundland. Ichnology is the study of the physical records of The beach on which the burrows were observed is behaviour, including, for example, tracks, trails, bur- approximately 90 m east of the mouth of Fifteen Mile rows, and borings (Bromley 1990). Studies are under- Brook (Figure 1). At the time of study, the beach was taken both in fossil and Recent contexts (palichnol- approximately 3 m wide by 24 m long. It passed on ogy and neoichnology, respectively). Fossilised traces the landward side into forest-covered glacio-fluvial are referred to as “trace fossils” or “ichnofossils” and sand and gravel (Figure 1), from which it was prob- are classified using Linnean binominal nomenclature. ably, in part, derived (cf., Batterson and Vatcher 1991). Although much ichnological research is focused on the Small driftwood logs were scattered over the beach. fossil record, neoichnology can provide crucial data The beach surface consisted of damp to wet (but not when interpreting ancient behaviours (e.g., Schäfer water-saturated), muddy to silty, fine- to medium- 1972; Chamberlain 1975; Ratcliffe and Fagerstrom grained sand containing local concentrations of finely 1980; Metz 1987). -

Official Journal L 338 Volume 35 of the European Communities 23 November 1992
ISSN 0378 - 6978 Official Journal L 338 Volume 35 of the European Communities 23 November 1992 English edition Legislation Contents I Acts whose publication is obligatory II Acts whose publication is not obligatory Council Council Directive 92 /92/ EEC of 9 November 1992 amending Directive 86/ 465 / EEC concerning the Community list of less-favoured farming areas within the meaning of Directive 75 / 268 / EEC (Federal Republic of Germany) 'New Lander* 1 Council Directive 92 /93 / EEC of 9 November 1992 amending Directive 75 /275 / EEC concerning the Community list of less-favoured farming areas within the meaning of Directive 75 / 268/ EEC (Netherlands) 40 Council Directive 92 / 94/ EEC of 9 November 1992 amending Directive 75 / 273 /EEC concerning the Community list of less-favoured farming areas within the meaning of Directive 75 / 268/ EEC (Italy) 42 2 Acts whose titles are printed in light type are those relating to day-to-day management of agricultural matters, and are generally valid for a limited period . The titles of all other Acts are printed in bold type and preceded by an asterisk. 23 . 11 . 92 Official Journal of the European Communities No L 338 / 1 II (Acts whose publication is not obligatory) COUNCIL COUNCIL DIRECTIVE 92/92/ EEC of 9 November 1992 amending Directive 86 /465 / EEC concerning the Community list of less-favoured farming areas within the meaning of Directive 75 /268 / EEC (Federal Republic of Germany ) 'New Lander' THE COUNCIL OF THE EUROPEAN COMMUNITIES , Commission of the areas considered eligible for inclusion -

Die Neue Kurparkbrücke Über Die Schwarza in Bad Blankenburg Steht
Gemeinsames Amts- und Mitteilungsblatt des Landkreises Saalfeld-Rudolstadt, der Städte Saalfeld/Saale, Rudolstadt und Bad Blankenburg 21/20 Amtsblatt: 27. JAHRGANG 26. November 2020 Während die Saalfelder Kranfirma GuR Saalfeld GmbH ihren Kran aufstellte, testeten Landrat und Bürgermeister als Erste die Brücke, die noch auf dem Transporter der Vorarlberger Transportfirma Vögel lagerte. Schließlich freuten sich Bürgermeister und die Hauptakteure über das gelun- gene Ergebnis, das als erste Zuschauer die Bärengruppe des Fröbel-Kindergartens bewunderte – insbesondere den Kran auf der anderen Uferseite der Schwarza. (Fotos: Martin Modes) Die neue Kurparkbrücke über die Schwarza in Bad Blankenburg steht Kurstadt-Verein hat ein wichtiges Ziel erreicht – Aluminium-Brückenüberbau aus Vorarlberg angeliefert Bad Blankenburg (AB/mmod). der Kurstadt-Verein mit seinen Fundamente und die Widerlager zu viele Interessierte dabei sein, „Das ist gut angelegtes Geld für Mitgliedern und Spenden von von der Firma Torus GmbH in die größtenteils vom gegenüber- die Bürgerinnen und Bürger der Bürgern und Firmen steht. Dazu Königsee haben einen Auftrags- liegenden Ufer verfolgten, wie die Stadt Bad Blankenburg – und gehört auch der Landkreis, der umfang von 115.000 Euro, der Brücke eingesetzt wurde. Eine auch für alle anderen Landkreis- den Kurstadt-Verein mit 25.000 Aluminium-Brückenüberbau, der richtige Einweihung mit den Bür- bürger, die den Kurpark besuchen Euro unterstützt. von der Bitschnau Metallverar- gern werde es hoffentlich Anfang wollen“, freut sich Landrat Marko Die Vereinsmitglieder hatten beitung GmbH aus dem österrei- Dezember noch geben, stellte der Wolfram über die neue Kurpark- seit Jahren um Unterstützung chischen Nenzing in Vorarlberg Bürgermeister in Aussicht. brücke, die am 12. November für den Wiederaufbau der 2013 angefertigt wurde, hat ein Kosten- Bereits um 10 Uhr am Vormit- eingesetzt wurde. -

Hierarchical Random Walks in Trace Fossils and the Origin of Optimal Search Behavior
Hierarchical random walks in trace fossils and the origin of optimal search behavior David W. Simsa,b,c,1, Andrew M. Reynoldsd, Nicolas E. Humphriesa, Emily J. Southalla, Victoria J. Wearmoutha, Brett Metcalfee, and Richard J. Twitchettf aMarine Biological Association of the United Kingdom, Plymouth PL1 2PB, United Kingdom; bOcean and Earth Science, National Oceanography Centre Southampton, University of Southampton, Waterfront Campus, Southampton SO14 3ZH, United Kingdom; cCentre for Biological Sciences, University of Southampton, Highfield Campus, Southampton SO17 1BJ, United Kingdom; dRothamsted Research, Harpenden, Hertfordshire AL5 2JQ, United Kingdom; eEarth and Climate Cluster, Faculty of Earth and Life Sciences, VU University Amsterdam, 1081 HV, Amsterdam, The Netherlands; and fDepartment of Earth Sciences, The Natural History Museum, London SW7 5BD, United Kingdom Edited by H. Eugene Stanley, Boston University, Boston, MA, and approved June 19, 2014 (received for review April 1, 2014) −μ Efficient searching is crucial for timely location of food and other “walk clusters” with no characteristic scale, such that P(l) ∼ l , resources. Recent studies show that diverse living animals use a with 1 < μ ≤ 3, where l is the move step length between turns and theoretically optimal scale-free random search for sparse resources μ the power-law exponent. Over many iterations, a Lévy walk will known as a Lévy walk, but little is known of the origins and evo- be distributed much further from its starting position than a lution of foraging behavior and the search strategies of extinct Brownian walk of the same length [hence is termed super- organisms. Here, using simulations of self-avoiding trace fossil trails, diffusive (8)], because small-step walk clusters are interspersed we show that randomly introduced strophotaxis (U-turns)—initiated by long “steps” to new locations, with this pattern repeating by obstructions such as self-trail avoidance or innate cueing—leads across all scales. -

Amtsblatt 01/2020 Vom 23. Januar 2020
Gemeinsames Amts- und Mitteilungsblatt des Landkreises Saalfeld Rudolstadt, der Städte Saalfeld/Saale, Rudolstadt und Bad Blankenburg 01/20 Amtsblatt: 27. JAHRGANG 23. Januar 2020 Simon Michalowicz zeigt Bilder von seiner 3.000-Kilometer-Fuß- Unter Bären in Alaska hat sich der Biologe und Bärenexperte David tour durch Norwegen. (Foto: Simon Michalowicz) Bittner begeben und stellt seine Erlebnisse vor. (Foto: David Bittner) Pilgern – Wege der Stille ist der Titel des Vortrags von Dieter Glogow- Im Oldtimer fuhren Sabine Hoppe und Thomas Rahn 30.000 Kilome- ski, Andrea Nuss und Stefan Rosenboom. (Foto: Dieter Glogowski) ter durch das wilde Südamerika. (Foto: Thomas Rahn) 22. Thüringer Weltsichen Festival in der Stadthalle Bad Blankenburg Vom 31. Januar bis zum 2. Februar zeigen erfahrene Reisende und Fotografen ihre Eindrücke der Welt Bad Blankenburg (AB/pl). Noch Smog, Stress und Lärm fordern Hintergründe und körperliche nie befand sich die Welt in einem ihren Tribut und so besinnen Herausforderungen, aber natür- solchen Wandel wie heute. Wo sich immer mehr Menschen auf lich auch die Welt im Wandel der sich noch vor kurzem endlose andere, tiefere Werte und bege- Zeit – inklusive einer Reise zu Regenwälder erstreckten, wach- ben sich in der Freizeit auf Pil- Völkern, die diesen Wandel kaum sen heute gigantische Ölpalmen- gerpfade. erleben – dies alles stellen erfah- plantagen. Staubige Dreckpisten Andere reisen in die entlegensten rene Reisende und großartige von einst sind längst zu dicht Ecken der Erde oder suchen ex- Fotografen vom 31. Januar bis befahrenen Autobahnen mutiert. treme Herausforderungen. Diese zum 2. Februar in der Stadthalle Lebten noch vor wenigen Jahren Vielfalt versuchen wir mit dem Bad Blankenburg in eindrucks- große Bevölkerungsteile auf dem 22. -

The Discovery of a Paleodictyon-Like Trace Fossil from the Late Cambrian Machinchang Formation in Pulau Jemuruk, Langkawi, Malaysia
Geological Society of Malaysia, Bulletin 46 May 2003; pp. 421-424 The discovery of a Paleodictyon-like trace fossil from the Late Cambrian Machinchang Formation in Pulau Jemuruk, Langkawi, Malaysia MOHD SHAFEEA LEMAN School of Environmental Sciences and Natural Resources, Faculty of Science and Technology Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Abstract: Closely spaced polygonal Paleodictyon-like trace fossil was recently discovered in Late Cambrian Machinchang Formation at Pulau Jemuruk, Langkawi. Each polygon is bounded by six segments of ridges forming unique hexagonal framework. The Jemuruk fossil has an epi-relief origin, contrary to the hypo-relief burrow Paleodictyon. The Jemuruk trace fossils formed in relatively shallower marine environment compared with other known Paleodictyon. This study shows that the Kinneyia structure found in the same layer originated from these Paleodictyon-like trace fossils. Abstrak: Surihan berbentuk poligon menyerupai Paleodictyon telah ditemui dalam batuan Kambria Akhir Formasi Machinchang di Pulau Jemuruk, Langkawi. Setiap ruang fosil surih ini disempadani enam segmen permatang membentuk kerangka heksagon yang menarik. Fosil Jemuruk berasalan epi-jasatimbul manakala Paleodictyon adalah sejenis korekan hipo-jasatimbul. Fosil surih Jemuruk terbentuk pada sekitaran laut lebih cetek berbanding dengan Paleodictyon yang diketahui. Kajian juga menunjukkan bahawa struktur Kinneyia yang ditemui dalam lapisan yang sarna adalah berasal daripada fosil surih seakan Paleodictyon ini. INTRODUCTION the Tarutao Formation described (later) by Teroaka et al. (1982) from Ko Tarutao in southern Thailand. The later The Machinchang Formation is perhaps the best has a comparatively similar lithological sequence with the described Cambrian rock formation in Malaysia. Despite Machinchang Formation and is a northern extension of the being one of the oldest rock formation in the country, most Machinchang Formation beyond the Malaysia-Thailand of its rocks are neither strongly deformed nor highly border. -
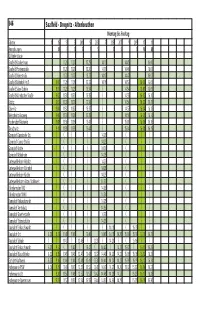
546 Saalfeld - Drognitz - Altenbeuthen Montag Bis Freitag Fahrtnr
546 Saalfeld - Drognitz - Altenbeuthen Montag bis Freitag Fahrtnr. 1 15 13 3 201 5 203 7 205 207 9 209 17 11 Anmerkungen M F S S F S S S S M 1 Gültigkeitstage Saalfeld Krankenhaus 11. 20 11. 20 13.25 14.10 14.45 16.00 Saalfeld Pfortenstraße 11. 21 11. 21 13.26 14.11 14.46 16.01 Saalfeld Dürerstraße 11. 22 11. 22 13.27 14.12 14.47 16.02 Saalfeld Bahnhof Hst.5 9.00 11. 28 11. 28 13.33 14.18 14.53 16.00 16.08 Saalfeld Sabel-Schule 9.01 11. 29 11. 29 13.34 {_ 14.54 16.01 16.09 Saalfeld Kulmbacher Straße 9.02 11. 3 0 11. 3 0 13.35 {_ 14.55 16.02 16.10 Köditz 9.03 11. 31 11. 31 13.36 {_ 14.56 16.03 16.11 Obernitz 9.04 11. 32 11. 32 13.37 {_ 14.57 16.04 16.12 Weischwitz Abzweig 9.06 11. 34 11. 34 13.39 {_ 14.59 16.05 16.14 Fischersdorf Abzweig 9.08 11. 3 6 11. 3 6 13.41 {_ 15.01 16.06 16.16 Tauschwitz 9.11 11. 39 11. 39 13.44 {_ 15.04 16.08 16.19 Gorndorf Gorndorfer Str. {_ {_ {_ {_ 14.21 {_ {_ {_ Gorndorf Geraer Straße {_ {_ {_ {_ 14.23 {_ {_ {_ Gorndorf Kirche {_ {_ {_ {_ 14.24 {_ {_ {_ Gorndorf Wohnheim {_ {_ {_ {_ 14.25 {_ {_ {_ Unterwellenborn Röblitz {_ {_ {_ {_ 14.27 {_ {_ {_ Unterwellenborn Bahnhof {_ {_ {_ {_ 14.28 {_ {_ {_ Unterwellenborn Kirche {_ {_ {_ {_ 14.29 {_ {_ {_ Unterwellenborn Abzw. -

Regionales Flächenmanagement Im Städtedreieck Am Saalebogen – Konzeption, Handhabe Und Weiterentwicklung –
Technische Universität Dresden Fakultät für Architektur Institut für Städtebau und Regionalplanung Dissertation zum Thema: Regionales Flächenmanagement im Städtedreieck am Saalebogen – Konzeption, Handhabe und Weiterentwicklung – Vorgelegt von: Robert Koch Referenten: Prof. Dr. Rainer Winkel Prof. Dr. Rainer Danielzyk Prof. Dr. Gerold Kind Verzeichnisse i Inhaltsverzeichnis Tabellenverzeichnis................................................................................................................................iv Abbildungsverzeichnis.............................................................................................................................v Abkürzungsverzeichnis...........................................................................................................................vi Vorwort und Danksagung.......................................................................................................................ix 1 EINFÜHRUNG: THEMATIK, ZIELSETZUNG UND VORGEHENSWEISE........................................................................................................1 2 DAS SELBSTVERSTÄNDNIS DER RAUMPLANUNG: ZWISCHEN PLANUNG UND ENTWICKLUNG ...............................................................................................................9 2.1 DIE ENTWICKLUNG EINER UMFASSENDEN RAUMPLANUNG IN DEUTSCHLAND ................10 2.2 AKTUELLE RAHMENBEDINGUNGEN UND REAKTION DER RAUMPLANUNG DARAUF ..........14 2.3 EINE ‚NEUE’ PLANUNGSKULTUR..................................................................................17 -

Cambrian Substrate Revolution
Vol. 10, No. 9 September 2000 INSIDE • Research Grants, p. 12 • Section Meetings Northeastern, p. 16 GSA TODAY Southeastern, p. 18 A Publication of the Geological Society of America • Happy Birthday, NSF, p. 22 The Cambrian Substrate Revolution David J. Bottjer, Department of Earth Sciences, University of Southern California, Los Angeles, CA 90089-0740, [email protected] James W. Hagadorn, Division of Geological and Planetary Sciences, California Institute of Technology, Pasadena, CA 91125, [email protected] Stephen Q. Dornbos, Department of Earth Sciences, University of Southern California, Los Angeles, CA 90089-0740, [email protected] ABSTRACT The broad marine ecological settings prevalent during the late Neo- proterozoic–early Phanerozoic (600–500 Ma) interval of early metazoan body plan origination strongly impacted the subsequent evolution and development of benthic metazoans. Recent work demonstrates that late Neoproterozoic seafloor sediment had well-developed microbial mats and poorly developed, vertically oriented bioturbation, thus producing fairly stable, relatively low water content substrates and a sharp water-sediment interface. Later in the Cambrian, seafloors with microbial mats became increasingly scarce in shallow-marine environments, largely due to the evolution of burrowing organisms with an increasing vertically oriented component to their bioturba- tion. The evolutionary and ecological effects of these substrate changes on Figure 1. Looping and meandering trace fossil Taphrhelminthopsis, made by a large Early Cambrian benthic metazoans, referred to as the bioturbator, on a bedding plane from Lower Cambrian Poleta Formation, White-Inyo Mountains, California. Such traces, consisting of a central trough between lateral ridges, occur in sandstones Cambrian substrate revolution, are deposited in shallow-marine environments. -

10 Ekdale & Gibert.Indd
SPANISH JOURNAL OF PALAEONTOLOGY Late Miocene deep-sea trace fossil associations in the Vera Basin, Almería, Southeastern Spain Allan A. EKDALE1* & Jordi M. de GIBERT2 1 Department of Geology and Geophysics, University of Utah, FASB 383, 115 South 1460 East, Salt Lake City, UT 84112- 0102, U.S.A.; [email protected] 2 Deceased (September 23, 2012) * Corresponding author Ekdale, A. & Gibert, J.M. 2014. Late Miocene deep-sea trace fossil associations in the Vera Basin, Almería, Southeastern Spain. [Asociaciones de trazas fósiles marinas profundas del Mioceno superior de la cuenca de Vera, Almería, Sureste de España]. Spanish Journal of Palaeontology, 29 (1), 95-104. Manuscript received 02 May 2013 © Sociedad Española de Paleontología ISSN 2255-0550 Manuscript accepted 12 September 2013 ABSTRACT RESUMEN The Vera Basin in southeastern Spain was a small, tectonically La Cuenca de Vera, en el sureste español, fue un pequeño active depocenter throughout the Miocene. In the early depocentro tectónicamente activo durante todo el Mioceno. Messinian, approximately 7.2 to 6.0 million years ago, A principios del Messiniense, hace aproximadamente 7,2- the basin received hemipelagic marl deposits that were 6,0 millones de años, la cuenca recibió depósitos margosos punctuated by turbidite events. Soles of thin, turbidite sand hemipelágicos, interrumpidos por episodios turbidíticos. beds preserve an abundance of pre-depositional graphoglyptid En la base de los niveles de arenas turbidíticas se conserva (agrichnial) burrows that represent diverse deep-sea una gran cantidad de grafoglíptidos pre-deposicionales ichnocoenoses, including Paleodictyon, Urohelminthoida (agrichnia) que representan diversas icnocenosis de aguas and Helminthorhaphe. Post-depositional feeding burrows, profundas, incluyendo Paleodictyon, Urohelminthoida including Ophiomorpha (created by crustaceans) and y Helminthorhaphe. -

Deep-Sea Trace Fossils in the West Crocker Formation, Sabah (Malaysia), and Their Palaeoenvironmental Significance Mazlan Madon
Bulletin of the Geological Society of Malaysia, Volume 71, May 2021, pp. 23 - 46 DOI: https://doi.org/10.7186/bgsm71202103 Deep-sea trace fossils in the West Crocker Formation, Sabah (Malaysia), and their palaeoenvironmental significance Mazlan Madon Malaysian Continental Shelf Project, National Security Council, Kuala Lumpur, Malaysia Author email address: [email protected] Abstract: In the “flysch” series of the West Crocker Formation (Eocene–Oligocene), Kota Kinabalu, Sabah, trace fossils are fairly common although not ubiquitous. The trace fossils commonly occur as hypichnial semi- or full-reliefs on the sole of thin turbiditic sandstone beds (mainly Bouma Tc division) in the thinly bedded heterolithic sandstone-mudstone facies interpreted as submarine fan lobe deposits. Their presence in mainly the thinly bedded facies of the fan system suggests preferential production and preservation in the fine-grained “distal” parts of the Crocker submarine fan system. Trace fossil assemblages characteristic of the Nereites ichnofacies indicate sedimentary environments mainly in bathyal to abyssal water depths (>2000 m). This ichnofacies is dominated by horizontal grazing, farming and feeding traces, ranging from solitary to branching tubular burrows (Ophiomorpha, Palaeophycus and Planolites) to meandering trails and tunnels (Nereites, Cosmorhaphe, Helminthopsis), as well as the spiriform burrows Spirophycus. Graphoglyptids are the most diagnostic of the Nereites ichnofacies, produced by sediment grazers and farmers (agrichnia) and often displaying intricate networks of mainly horizontal tunnels preserved as hypichnial semi-reliefs. They include the delicate spiral traces of Spirorhaphe, as well as the enigmatic hexagonal network burrow Paleodictyon. Other ichnogenera include Planolites, Thalassinoides and Ophiomorpha which are facies-crossing and not environment specific.