The Architecture of the Baby Brain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans Ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai
Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai To cite this version: Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai. Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex. Frontiers in Neuroanatomy, Frontiers, 2018, 12, pp.93. 10.3389/fnana.2018.00093. hal-01929541 HAL Id: hal-01929541 https://hal.archives-ouvertes.fr/hal-01929541 Submitted on 21 Nov 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. REVIEW published: 19 November 2018 doi: 10.3389/fnana.2018.00093 Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans J. ten Donkelaar 1*†, Nathalie Tzourio-Mazoyer 2† and Jürgen K. Mai 3† 1 Department of Neurology, Donders Center for Medical Neuroscience, Radboud University Medical Center, Nijmegen, Netherlands, 2 IMN Institut des Maladies Neurodégénératives UMR 5293, Université de Bordeaux, Bordeaux, France, 3 Institute for Anatomy, Heinrich Heine University, Düsseldorf, Germany The gyri and sulci of the human brain were defined by pioneers such as Louis-Pierre Gratiolet and Alexander Ecker, and extensified by, among others, Dejerine (1895) and von Economo and Koskinas (1925). -

Acetylcholinesterase Staining in Human Auditory and Language
Acetylcholinesterase Staining in Human Jeffrey J. Hutsler and Michael S. Gazzaniga Auditory and Language Cortices: Center for Neuroscience, University of California, Davis Regional Variation of Structural Features California 95616 Cholinergic innovation of the cerebral neocortex arises from the basal 1974; Greenfield, 1984, 1991; Robertson, 1987; Taylor et al., forebrain and projects to all cortical regions. Acetylcholinesterase 1987; Krisst, 1989; Small, 1989, 1990). AChE-containing axons (AChE), the enzyme responsible for deactivating acetylcholine, is found of the cerebral cortex are also immunoreactive for choline within both cholinergic axons arising from the basal forebrain and a acetyltransferase (ChAT) and are therefore known to be cho- Downloaded from subgroup of pyramidal cells in layers III and V of the cerebral cortex. linergic (Mesulam and Geula, 1992). This pattern of staining varies with cortical location and may contrib- AChE-containing pyramidal cells of layers III and V are not ute uniquely to cortical microcircuhry within functionally distinct cholinergic (Mesulam and Geula, 1991), but it has been sug- regions. To explore this issue further, we examined the pattern of AChE gested that they are cholinoceptive (Krnjevic and Silver, 1965; staining within auditory, auditory association, and putative language Levey et al., 1984; Mesulam et al., 1984b). In support of this regions of whole, postmortem human brains. notion, layer in and V pyramidal cell excitability can be mod- http://cercor.oxfordjournals.org/ The density and distribution of acetylcholine-containing axons and ulated by the application of acetylcholine in the slice prepa- pyramidal cells vary systematically as a function of auditory process- ration (McCormick and Williamson, 1989). Additionally, mus- ing level. -
Review of Temporal Lobe Structures
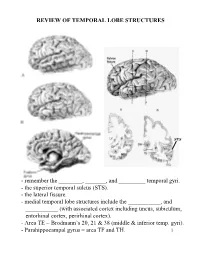
REVIEW OF TEMPORAL LOBE STRUCTURES STS - remember the ________, _______, and _________ temporal gyri. - the superior temporal sulcus (STS). - the lateral fissure. - medial temporal lobe structures include the ___________, and ___________ (with associated cortex including uncus, subiculum, entorhinal cortex, perirhinal cortex). - Area TE = Brodmann’s 20, 21 & 38 (middle & inferior temp. gyri). - Parahippocampal gyrus = area TF and TH. 1 TEMPORAL LOBE FUNCTIONS Sensory Inputs to Temporal lobe: 1. ____________________________________________; 2. _________________________________________________. Temporal cortical regions and functional correlates: 1. Within lateral fissure (superior surface of lateral fissure): a) Heschel’s gyri (_____________________). b) posterior to Heschel’s gyri (_______________________). c) Planum temporale (secondary auditory cortex; Wernicke’s area - specialized in __________________________). 2 Temporal cortical regions and functional correlates (continued): 2. Superior temporal sulcus, middle and inferior temporal gyrus (area TE): _________________________________________ ____________. 3. Ventral/medial surface of temporal lobe (hippocampus and associated cortex): ______________________________. - the ventral/medial surface of the temporal lobe is also associated with the amygdala. Together with the surrounding ventral/medial temporal lobe, the amygdala is involved in __________________________________________. Hemispheric “specialization”: 1. Left hemisphere: a) ________________; b) ____________________________. -

Surgical Anatomy and Techniques
SURGICAL ANATOMY AND TECHNIQUES MICROSURGICAL APPROACHES TO THE MEDIAL TEMPORAL REGION:AN ANATOMICAL STUDY Alvaro Campero, M.D. OBJECTIVE: To describe the surgical anatomy of the anterior, middle, and posterior Department of Neurological Surgery, portions of the medial temporal region and to present an anatomic-based classification University of Florida, of the approaches to this area. Gainesville, Florida METHODS: Twenty formalin-fixed, adult cadaveric specimens were studied. Ten brains Gustavo Tro´ccoli, M.D. provided measurements to compare different surgical strategies. Approaches were demon- Department of Neurological Surgery, strated using 10 silicon-injected cadaveric heads. Surgical cases were used to illustrate the Hospital “Dr. J. Penna,” results by the different approaches. Transverse lines at the level of the inferior choroidal point Bahı´a Blanca, Argentina and quadrigeminal plate were used to divide the medial temporal region into anterior, middle, and posterior portions. Surgical approaches to the medial temporal region were classified into Carolina Martins, M.D. four groups: superior, lateral, basal, and medial, based on the surface of the lobe through which Department of Neurological Surgery, University of Florida, the approach was directed. The approaches through the medial group were subdivided further Gainesville, Florida into an anterior approach, the transsylvian transcisternal approach, and two posterior ap- proaches, the occipital interhemispheric and supracerebellar transtentorial approaches. Juan C. Fernandez-Miranda, M.D. RESULTS: The anterior portion of the medial temporal region can be reached through Department of Neurological Surgery, University of Florida, the superior, lateral, and basal surfaces of the lobe and the anterior variant of the Gainesville, Florida approach through the medial surface. -

The Depth Asymmetry of Superior Temporal Sulcus
New human-specific brain landmark: The depth asymmetry of superior temporal sulcus François Leroya,1, Qing Caib, Stephanie L. Bogartc, Jessica Duboisa, Olivier Coulond, Karla Monzalvoa, Clara Fischere, Hervé Glasela, Lise Van der Haegenf, Audrey Bénézita, Ching-Po Ling, David N. Kennedyh, Aya S. Iharai, Lucie Hertz-Pannierj, Marie-Laure Moutardk, Cyril Pouponl, Marc Brysbaerte, Neil Robertsm, William D. Hopkinsc, Jean-François Mangine, and Ghislaine Dehaene-Lambertza aCognitive Neuroimaging Unit, U992, eAnalysis and Processing of Information Unit, jClinical and Translational Applications Research Unit, U663, and lNuclear Magnetic Resonance Imaging and Spectroscopy Unit, Office of Atomic Energy and Alternative Energies (CEA), INSERM, NeuroSpin, Gif-sur-Yvette 91191, France; bShanghai Key Laboratory of Brain Functional Genomics, East China Normal University, Shanghai 200241, China; cDivision of Developmental and Cognitive Neuroscience, Yerkes National Primate Research Center, Atlanta, GA 30322; dLaboratory for Systems and Information Science, UMR CNRS 7296, Aix-Marseille University, Marseille 13284, France; fDepartment of Experimental Psychology, Ghent University, Ghent B-9000, Belgium; gInstitute of Neuroscience, National Yang-Ming University, Taipei City 112, Taiwan; hCenter for Morphometric Analysis, Neuroscience Center, Massachusetts General Hospital, Boston, MA 02114; iCenter for Information and Neural Networks, National Institute of Information and Communications Technology, Osaka 565-0871 Japan; kNeuropediatrics Department, Trousseau -

Pars Triangularis Asymmetry and Language Dominance ANNE L
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 719-722, January 1996 Neurobiology Pars triangularis asymmetry and language dominance ANNE L. FOUNDAS*, CHRISTIANA M. LEONARDO§, ROBIN L. GILMOREt¶, EILEEN B. FENNELLtII, AND KENNETH M. HEILMAN4:¶** *Department of Psychiatry and Neurology, Tulane University School of Medicine and Neurology Service, Veterans Affairs Medical Center, 1430 Tulane Avenue, New Orleans, LA 70112-2632; and *Center for Neuropsychological Studies, Departments of §Neuroscience and 1Neurology and I'Clinical and Health Psychology, University of Florida College of Medicine and **Neurology Service, Veterans Affairs Medical Center, Gainesville, FL 32608-1197 Communicated by Philip Teitelbaum, University of Florida, Gainesville, FL, October 10, 1995 (received for review March 15, 1995) ABSTRACT The pars triangularis is a portion of Broca's planum temporale can be performed (19-22). Given these area. The convolutions that form the inferior and caudal advances in MRI technology, reliable measures of language extent of the pars triangularis include the anterior horizontal laterality and handedness can be evaluated in the same indi- and anterior ascending rami of the sylvian fissure, respec- viduals, thus allowing direct structure-function correlations. tively. To learn if there are anatomic asymmetries of the pars Using this technique, Steinmetz et al. (22) studied planum triangularis, these convolutions were measured on volumetric temporale asymmetries in a group of left- and right-handers. magnetic resonance imaging scans of 11 patients who had A leftward asymmetry of the planum temporale was docu- undergone selective hemispheric anesthesia (Wada testing) to mented with a greater degree of leftward asymmetry in the determine hemispheric speech and language lateralization. -

Sex-Specific Differences in Resting-State Functional Connectivity
www.nature.com/scientificreports OPEN Sex‑specifc diferences in resting‑state functional connectivity of large‑scale networks in postconcussion syndrome Reema Shaf1,2*, Adrian P. Crawley3,4, Maria Carmela Tartaglia5,6,7,8, Charles H. Tator4,6,7,9,10, Robin E. Green1,2,3,4,6, David J. Mikulis3,4,6 & Angela Colantonio1,2,11 Concussions are associated with a range of cognitive, neuropsychological and behavioral sequelae that, at times, persist beyond typical recovery times and are referred to as postconcussion syndrome (PCS). There is growing support that concussion can disrupt network‑based connectivity post‑injury. To date, a signifcant knowledge gap remains regarding the sex‑specifc impact of concussion on resting state functional connectivity (rs‑FC). The aims of this study were to (1) investigate the injury‑ based rs‑FC diferences across three large‑scale neural networks and (2) explore the sex‑specifc impact of injury on network‑based connectivity. MRI data was collected from a sample of 80 concussed participants who fulflled the criteria for postconcussion syndrome and 31 control participants who did not have any history of concussion. Connectivity maps between network nodes and brain regions were used to assess connectivity using the Functional Connectivity (CONN) toolbox. Network based statistics showed that concussed participants were signifcantly diferent from healthy controls across both salience and fronto‑parietal network nodes. More specifcally, distinct subnetwork components were identifed in the concussed sample, with hyperconnected frontal nodes and hypoconnected posterior nodes across both the salience and fronto‑parietal networks, when compared to the healthy controls. Node‑to‑region analyses showed sex‑specifc diferences across association cortices, however, driven by distinct networks. -

'Absolute Pitch—Functional Evidence of Speech-Relevant Auditory Acuity'
Oechslin, M; Meyer, M; Jäncke, L (2010). Absolute Pitch—Functional Evidence of Speech-Relevant Auditory Acuity. Cordical Cortex, 20(2):447-455. Postprint available at: http://www.zora.uzh.ch University of Zurich Posted at the Zurich Open Repository and Archive, University of Zurich. Zurich Open Repository and Archive http://www.zora.uzh.ch Originally published at: Cordical Cortex 2010, 20(2):447-455. Winterthurerstr. 190 CH-8057 Zurich http://www.zora.uzh.ch Year: 2010 Absolute Pitch—Functional Evidence of Speech-Relevant Auditory Acuity Oechslin, M; Meyer, M; Jäncke, L Oechslin, M; Meyer, M; Jäncke, L (2010). Absolute Pitch—Functional Evidence of Speech-Relevant Auditory Acuity. Cordical Cortex, 20(2):447-455. Postprint available at: http://www.zora.uzh.ch Posted at the Zurich Open Repository and Archive, University of Zurich. http://www.zora.uzh.ch Originally published at: Cordical Cortex 2010, 20(2):447-455. Cerebral Cortex February 2010;20:447--455 doi:10.1093/cercor/bhp113 Advance Access publication July 10, 2009 Absolute Pitch—Functional Evidence of Mathias S. Oechslin, Martin Meyer and Lutz Ja¨ncke Speech-Relevant Auditory Acuity Department of Neuropsychology, University of Zurich, CH-8050 Zurich, Switzerland Absolute pitch (AP) has been shown to be associated with Interestingly, memorization of musical pieces without referring morphological changes and neurophysiological adaptations in the to a notation is an important key issue of this approach which planum temporale, a cortical area involved in higher-order auditory particularly puts emphasis on auditory features, namely and speech perception processes. The direct link between speech discrimination and representation of pitch and timbre. -
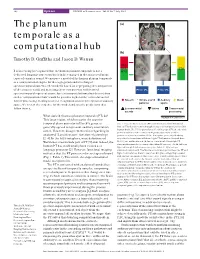
The Planum Temporale As a Computational
348 Opinion TRENDS in Neurosciences Vol.25 No.7 July 2002 The planum (a) temporale as a LR computational hub (b) Timothy D. Griffiths and Jason D. Warren STG, STS, STG It is increasingly recognized that the human planum temporale is not a MTG dedicated language processor, but is in fact engaged in the analysis of many types of complex sound. We propose a model of the human planum temporale as a computational engine for the segregation and matching of spectrotemporal patterns. The model is based on segregating the components of the acoustic world and matching these components with learned PTO / IPL PTO / IPL spectrotemporal representations. Spectrotemporal information derived from such a ‘computational hub’ would be gated to higher-order cortical areas for further processing, leading to object recognition and the perception of auditory Speech + Simple sound Auditory Music patterns space space. We review the evidence for the model and specific predictions that follow from it. Environmental Voices Cross-modal sounds processing What does the human planum temporale (PT) do? TRENDS in Neurosciences This large region, which occupies the superior temporal plane posterior to Heschl’s gyrus, is Fig. 1. The planum temporale (PT) as an anatomical and functional generally agreed to represent auditory association hub. (a) Tilted axial section through the superior temporal plane of the cortex. However, disagreement exists regarding its human brain. The PT lies posterior to Heschl’s gyrus (HG), the site of the primary auditory cortex, and is contiguous posteriorly with the anatomy [1] and structure–function relationships parieto–temporal operculum (PTO). Ninety-five percent probability [2–4]. -

A Surface-Based Analysis of Hemispheric Asymmetries and Folding of Cerebral Cortex in Term-Born Human Infants
2268 • The Journal of Neuroscience, February 10, 2010 • 30(6):2268–2276 Development/Plasticity/Repair A Surface-Based Analysis of Hemispheric Asymmetries and Folding of Cerebral Cortex in Term-Born Human Infants Jason Hill,1 Donna Dierker,2 Jeffrey Neil,1 Terrie Inder,1 Andrew Knutsen,3 John Harwell,2 Timothy Coalson,2 and David Van Essen2 Departments of 1Pediatrics, 2Anatomy and Neurobiology, and 3Mechanical, Aerospace, and Structural Engineering, Washington University School of Medicine, St. Louis, Missouri 63108 We have established a population average surface-based atlas of human cerebral cortex at term gestation and used it to compare infant and adult cortical shape characteristics. Accurate cortical surface reconstructions for each hemisphere of 12 healthy term gestation infants were generated from structural magnetic resonance imaging data using a novel segmentation algorithm. Each surface was inflated, flattened, mapped to a standard spherical configuration, and registered to a target atlas sphere that reflected shape character- isticsofall24contributinghemispheresusinglandmarkconstrainedsurfaceregistration.Populationaveragemapsofsulcaldepth,depth variability, three-dimensional positional variability, and hemispheric depth asymmetry were generated and compared with previously established maps of adult cortex. We found that cortical structure in term infants is similar to the adult in many respects, including the pattern of individual variability and the presence of statistically significant structural asymmetries in lateral temporal cortex, including the planum temporale and superior temporal sulcus. These results indicate that several features of cortical shape are minimally influ- enced by the postnatal environment. Introduction within the infant brain, it is of interest to determine whether the The adult human cerebral cortex is highly convoluted, variable features of regional variability and/or asymmetry that character- in shape across regions and across individuals, and asymmet- ize the adult cortex are already present at term. -

Redefining the Functional Organization of the Planum
Chapter 12 Redefi ning the Functional Organization of the Planum Temporale Region: Space, Objects, and Sensory–Motor Integration Gregory Hickok and Kourosh Saberi 12.1 Introduction: Defi nitions, History, and Preview The planum temporale (PT) is defi ned anatomically by the triangular surface on the supratemporal plane posterior to Heschl’s gyrus (see Clarke and Morosan, Chapter 2 ). Its posterior boundary is the termination point of the Sylvian fi ssure, its medial boundary is the insula or the point where the supratemporal plane transitions into the parietal operculum, and its lateral boundary is the lateral lip of the superior temporal gyrus (Fig. 12.1 ). Interest in the PT was promoted by the discovery of a left–right asymmetry in this structure, with the left PT being larger than the right in most (~65%) right-handed individuals (Geschwind & Levitsky, 1968 ) . Given that the left PT comprises a part of classical Wernicke’s area, the relatively larger size was thought to be an anatomical refl ection of the region’s functional specialization for speech processing. Several subsequent fi ndings cast doubt on this view. One was that PT asymmetry was found to correlate with a nonspeech function, musical ability (Schlaug et al., 1995 ) . Another was that a similar leftward PT asymmetry was found in chimpan- zees, a species without speech ability at all (Gannon et al., 1998 ; this of course questions the basis of the association with musical ability as well!). A third was that structural asymmetries of the PT did not correlate with language dominance as assessed directly using the intracarotid sodium amytal (Wada) procedure (Dorsaint- Pierre et al., 2006 ) . -

Supplemental Information Musical Expertise Shapes Functional And
Supplemental Information Musical expertise shapes functional and structural brain networks independent of absolute pitch ability Simon Leipold a, b, $, Carina Klein a, $, Lutz Jäncke a, c, d $ These authors contributed equally to this work a Division Neuropsychology, Department of Psychology, University of Zurich, Zurich, Switzerland b Department of Psychiatry and Behavioral Sciences, Stanford University School of Medicine, Stanford, USA c University Research Priority Program (URPP), Dynamics of Healthy Aging, University of Zurich, Zurich, Switzerland d Department of Special Education, King Abdulaziz University, Jeddah, Kingdom of Saudi Arabia 1 Supplemental Figures Supplementary Figure 1. Increased resting-state functional connectivity in AP musicians compared to non-musicians (pFWE < 0.05) in the seed- to-voxel analysis for the following seeds: A) the left HG, B) the right HG, C) the left PT and D) the right PT. Abbreviations: HG = Heschl’s gyrus; L = left; PT = planum temporale; R = right. 2 Supplementary Figure 2. Increased subnetworks in AP musicians compared to non-musicians obtained in the whole-brain NBS analysis for A) resting-state functional connectivity and B) DWI-based structural connectivity (pFWE < 0.05). Abbreviations: ACC = anterior cingulate cortex; aSMG = anterior supramarginal gyrus; aSTG = anterior superior temporal gyrus; cOp = central operculum; fOp = frontal operculum; FP = frontal pole; HG = Heschl’s gyrus; IFG,po = inferior frontal gyrus, pars opercularis; L = left; MTG = middle temporal gyrus; PCC = posterior cingulate cortex; postCG = postcentral gyrus; preCG = precentral gyrus; pSTG = superior temporal gyrus, posterior division; pOp = parietal operculum; PP = planum polare; PT = planum temporale; ptFG = posterior temporal fusiform gyrus; R = right; toFG = temporal occipital fusiform gyrus; TP = temporal pole.