Mesenchyme [1]
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Carl Gegenbaur – Wikipedia
Carl Gegenbaur – Wikipedia http://de.wikipedia.org/wiki/Carl_Gegenbaur aus Wikipedia, der freien Enzyklopädie Carl Gegenbaur (* 21. August 1826 in Würzburg; † 14. Juni 1903 in Heidelberg) war ein deutscher Arzt, einer der bedeutendsten Wirbeltiermorphologen des 19. Jahrhunderts und einer der Väter der Evolutionsmorphologie, eines modernen Begriffs von Walter J. Bock und Dwight D. Davis. 1 Leben 2 Wissenschaftliche Leistung 2.1 Das „Kopf-Problem“ 3 Verhältnis Carl Gegenbaur und Ernst Haeckel 4 Nach Gegenbaur benannte Taxa Carl Gegenbaur 5 Ausgewählte Arbeiten 6 Weiterführende Literatur 7 Weblinks 8 Einzelnachweise Er war der Sohn des Justizbeamten Franz Joseph Gegenbaur (1792–1872) und dessen Ehefrau Elisabeth Karoline (1800–1866), geb. Roth. Von seinen sieben Geschwistern verstarben vier sehr jung; sein um drei Jahre jüngerer Bruder wurde nur 25 Jahre und seine um 13 Jahre jüngere Schwester 38 Jahre alt.[1] 1838 besuchte er die Lateinschule und von 1838 bis 1845 in Würzburg das Gymnasium. Schon zu seiner Schulzeit führt er Naturstudien in der Umgebung von Würzburg und bei Verwandten im Odenwald (Amorbach) durch. Er beschäftigt sich weiterführend mit Pflanzen, Tieren, Gesteinen, legte Sammlungen an, fertigte Zeichnungen und führte erste Tiersektionen durch.[2] Politische Hintergründe: Bayern wurde 1806 unter dem Minister Maximilian Carl Joseph Franz de Paula Hieronymus Graf von Montgelas (1759–1838) zu einem der führenden Mitglieder im Rheinbund und Bündnispartner von Napoleon Bonaparte (1769–1821). Für seine Bündnistreue wurde im Frieden von Pressburg Bayern zum Königreich durch den französischen Kaiser aufgewertet und Herzog Maximilian IV. (1756–1825) am 1. Januar 1806 in München als Maximilian I. Joseph zum ersten König Bayerns erhoben. -

The Fingerprint Sourcebook
CHAPTER EMBRYOLOGY AND MORPHOLOGY OF FRICTION RIDGE SKIN Kasey Wertheim CONTENTS 3 3.1 Introduction 12 3.7 Pattern Formation 4 3.2 Embryology: Establishing 18 3.8 Genetics Uniqueness and Pattern Formation in the Friction Ridge Skin 5 3.3 Limb Development 21 3.9 Uniqueness: Developmental Noise 7 3.4 Differentiation of the Friction 22 3.10 Summary: Keys to Ridge Skin Uniqueness and Pattern Formation 8 3.5 Primary Ridge Formation 22 3.11 Reviewers 11 3.6 Secondary Ridge 24 3.12 References Formation 3–1 Embryology and Morphology of Friction Ridge Skin C H A P T E R 3 CHAPTER 3 EMBRYOLOGY AND 3.1 Introduction Friction ridge skin has unique features that persist from MORPHOLOGY OF before birth until decomposition after death. Upon contact with a surface, the unique features of friction ridge skin may leave an impression of corresponding unique details. Two FRICTION RIDGE SKIN impressions can be analyzed, compared, and evaluated, and if sufficient quality and quantity of detail is present (or Kasey Wertheim lacking) in a corresponding area of both impressions, a com- petent examiner can effect an individualization or exclusion (identify or exclude an individual). The analysis, comparison, evaluation, and verification (ACE-V) methodology, combined with the philosophy of quantitative–qualitative examinations, provide the framework for practical application of the friction ridge examination discipline. But at the heart of the disci- pline is the fundamental principle that allows for conclusive determinations: the source of the impression, friction ridge skin, is unique and persistent. Empirical data collected in the medical and forensic com- munities continues to validate the premises of uniqueness and persistence. -

Development of the Skin and Its Derivatives
Development of the skin and its derivatives Resources: http://php.med.unsw.edu.au/embryology/ Larsen’s Human Embryology – Chapter 7 The Developing Human: Clinically Oriented Embryology Dr Annemiek Beverdam – School of Medical Sciences, UNSW Wallace Wurth Building Room 234 – [email protected] Lecture overview Skin function and anatomy Skin origins Development of the overlying epidermis Development of epidermal appendages: Hair follicles Glands Nails Teeth Development of melanocytes Development of the Dermis Resources: http://php.med.unsw.edu.au/embryology/ Larsen’s Human Embryology – Chapter 7 The Developing Human: Clinically Oriented Embryology Dr Annemiek Beverdam – School of Medical Sciences, UNSW Wallace Wurth Building Room 234 – [email protected] Skin Function and Anatomy Largest organ of our body Protects inner body from outside world (pathogens, water, sun) Thermoregulation Diverse: thick vs thin skin, scalp skin vs face skin, etc Consists of: - Overlying epidermis - Epidermal appendages: - Hair follicles, - Glands: sebaceous, sweat, apocrine, mammary - Nails - Teeth - Melanocytes - (Merkel Cells - Langerhans cells) - Dermis - Hypodermis Skin origins Trilaminar embryo Ectoderm (Neural crest) brain, spinal cord, eyes, peripheral nervous system epidermis of skin and associated structures, melanocytes, cranial connective tissues (dermis) Mesoderm musculo-skeletal system, limbs connective tissue of skin and organs urogenital system, heart, blood cells Endoderm epithelial linings of gastrointestinal and respiratory tracts Ectoderm -
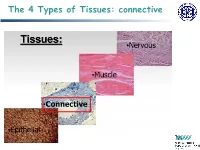
The 4 Types of Tissues: Connective
The 4 Types of Tissues: connective Connective Tissue General structure of CT cells are dispersed in a matrix matrix = a large amount of extracellular material produced by the CT cells and plays a major role in the functioning matrix component = ground substance often crisscrossed by protein fibers ground substance usually fluid, but it can also be mineralized and solid (bones) CTs = vast variety of forms, but typically 3 characteristic components: cells, large amounts of amorphous ground substance, and protein fibers. Connective Tissue GROUND SUBSTANCE In connective tissue, the ground substance is an amorphous gel-like substance surrounding the cells. In a tissue, cells are surrounded and supported by an extracellular matrix. Ground substance traditionally does not include fibers (collagen and elastic fibers), but does include all the other components of the extracellular matrix . The components of the ground substance vary depending on the tissue. Ground substance is primarily composed of water, glycosaminoglycans (most notably hyaluronan ), proteoglycans, and glycoproteins. Usually it is not visible on slides, because it is lost during the preparation process. Connective Tissue Functions of Connective Tissues Support and connect other tissues Protection (fibrous capsules and bones that protect delicate organs and, of course, the skeletal system). Transport of fluid, nutrients, waste, and chemical messengers is ensured by specialized fluid connective tissues, such as blood and lymph. Adipose cells store surplus energy in the form of fat and contribute to the thermal insulation of the body. Embryonic Connective Tissue All connective tissues derive from the mesodermal layer of the embryo . The first connective tissue to develop in the embryo is mesenchyme , the stem cell line from which all connective tissues are later derived. -

The Role of the Mesenchyme in 1. Patterns of Mesenchymal Cell
THE AMERICAN JOURNAL OF ANATOMY 188:121-132 (1990) The Role of the Mesenchyme in Mouse Neural Fold Elevation 1. Patterns of Mesenchymal Cell Distribution and Proliferation in Embryos Developing In Vitro JOYCE MORRIS-WIMAN AND LINDA L. BRINKLEY Department of Anatomy & Cell Biology, University of Michigan Medical School, Ann Arbor, Michigan 48109 ABSTRACT Using the computer-assisted a decrease in the density of cells residing in the central method of smoothed spatial averaging, spatial region of the mesenchyme and with a decrease in the and temporal patterns of cell distribution and mi- concentration of hyaluronate (HA) in this region (Mor- totic activity were analyzed in the cranial mesen- ris-Wiman and Brinkley, 1989). The decrease observed chyme underlying the mesencephalic neural folds in mesenchymal cell density during elevation could re- sult from a decrease in mitotic rate, from an increase in of mouse embryos maintained in roller tube cul- cell death, or from the expanion of the extracellular ture. Total cell density increased in central and matrix of the cranial mesenchyme displacing cells from medial mesenchymal regions after 12 hr in cul- this region. The glycosaminoglycan hyaluronate is the ture, decreased after 18 hr, and showed a further major component of this extracellular matrix (Solursh decrease after 24 hr when the neural folds of the and Morriss, 1977; Morriss and Solursh, 1978a,b;Hei- embryos had elevated, converged, and were fus- fetz et al., 1980). In biological and experimental sys- ing or fused. Mitotic activity, as measured by the tems, it is capable of a high degree of hydration with a ratio of 3H-thymidine-labeled cells to unlabeled concomitant increase in volume (Comper and Laurent, cells, was highest in the central mesenchyme at all 1978). -

The Dancing Bees: Karl Von Frisch, the Honeybee Dance Language
The Dancing Bees: Karl von Frisch, the Honeybee Dance Language, and the Sciences of Communication By Tania Munz, Research Fellow MPIWG, [email protected] In January of 1946, while much of Europe lay buried under the rubble of World War Two, the bee researcher Karl von Frisch penned a breathless letter from his country home in lower Austria. He reported to a fellow animal behaviorist his “sensational findings about the language of the bees.”1 Over the previous summer, he had discovered that the bees communicate to their hive mates the distance and direction of food sources by means of the “dances” they run upon returning from foraging flights. The straight part of the figure-eight-shaped waggle dance makes the same angle with the vertical axis of the hive as the bee’s flight line from the hive made with the sun during her outgoing flight. Moreover, he found that the frequency of individual turns correlated closely with the distance of the food; the closer the supply, the more rapidly the bee dances. Von Frisch’s assessment in the letter to his colleague would prove correct – news of the discovery was received as a sensation and quickly spread throughout Europe and abroad. In 1973, von Frisch was awarded the Nobel Prize in Physiology or Medicine together with the fellow animal behaviorists Konrad Lorenz and Niko Tinbergen. The Prize bestowed public recognition that non-human animals possess a symbolic means of communication. Dancing Bees is a dual intellectual biography – about the life and work of the experimental physiologist Karl von Frisch on the one hand and the honeybees as cultural, experimental, and especially communicating animals on the other. -

Role of Extracellular Matrix in Regulating Embryonic Epithelial-Mesenchymal Transition
BioMol Concepts, Vol. 3 (2012), pp. 333–344 • Copyright © by Walter de Gruyter • Berlin • Boston. DOI 10.1515/bmc-2011-0065 Review Role of extracellular matrix in regulating embryonic epithelial-mesenchymal transition Francesca Zito considerable amount of data about its basic mechanisms. Istituto di Biomedicina e Immunologia Molecolare From a clinical viewpoint, the results obtained so far offer ‘ Alberto Monroy ’ , Consiglio Nazionale delle Ricerche, a great promise for the development of new therapies. At Via Ugo La Malfa 153, I-90146 Palermo , Italy the same time, they advance our knowledge about essential developmental processes. e-mail: [email protected] Although EMT is a widely studied topic, it is likely that not all aspects of this process have been elucidated so far. Considerable knowledge has been accumulated regarding cell Abstract morphological changes, and the regulation of several genes and various signaling pathways have been partially under- Embryogenesis and morphogenesis are characterized by stood. Still, little is known about the role of the extracellular complex cell rearrangements and movements which require matrix (ECM) as a cell substrate during EMT, and cell motility appropriate interactions of cells with the surrounding extracel- is probably the least understood aspect of the whole process. lular matrix (ECM) by means of specifi c membrane receptors. Many events in developing embryo are associated with Interest in the identifi cation and purifi cation of ECM com- cell migration, which consists of the coordinated move- ponents, as well as in conducting functional studies of them, ments of several different cells over short or long distances including their ligands and other molecules involved in cell- throughout the embryo. -

Metamorphic Changes of Dermis in Skin of Frog Larvae Exposed to Thyroxine’
DEVELOPMENTAL BIOLOGY, 7, 244-254 (1963) Metamorphic Changes of Dermis in Skin of Frog Larvae Exposed to Thyroxine’ NORMAN E. KEMP Department of Zoology, The University of Michigan, Ann Arbor, Michigan Accepted October 22, 1962 INTRODUCTION Skin in the tadpole of anuran larvae prior to metamorphosis is composed of two layers of epidermal cells (Leeson and Threadgold, 1961; Chapman and Dawson, 1961; Edds and Sweeny, 1960, 1962), a laminated basement lamella, and a layer of dermal mesenchyme cells apposed to the underside of the basement lamella. An adepidermal space and membrane (Salpeter and Singer, 1959, 1960) separate epi- dermis from the dermal basement lamella. Weiss and Ferris (1954) first observed that mesenchyme cells migrate into the basement lamella as metamorphosis begins. Invasion of the lamella has been reported to begin about Taylor-Kollros stage XI or XII (Kemp, 1961b) and to proceed until mesenchyme cells move entirely through the lamella and cause it to become detached from the adepidermal mem- brane at stages XV-XVI. Subsequently a population of cells consti- tuting the stratum spongiosum develops beneath the adepiderma1 membrane and the basement lamella becomes the deeper-lying stra- tum compacturn or “dermal lamella” of the adult dermis. New poly- merization of collagenous fibers and ground substance results in deposition of the adult basement membrane between the adepidermal membrane and the stratum spongiosum. Generalized metamorphic changes can be induced in amphibian larvae by exposing them to thyroxine or other thyroid hormones (Kollros, 1959; Pitt-Rivers and Tata, 1959). Local metamorphic changes can be induced by implanting pellets of thyroxine mixed with ‘This investigation was supported by grants from the United States Public Health Service (RG 5867-C>), the National Science Foundation (G-19447), and the Michigan Memorial-Phoenix Project (no. -

Preface for Uchida Anniversary Volume
Title PREFACE FOR UCHIDA ANNIVERSARY VOLUME Author(s) WITSCHI, Emil Citation 北海道大學理學部紀要, 13(1-4) Issue Date 1957-08 Doc URL http://hdl.handle.net/2115/27187 Type bulletin (editorial) File Information 13(1_4)_Preface.pdf Instructions for use Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP 'V PREFACE FOR UCHIDA ANNIVERSARY VOLUME My personal acquaintance with Tohru Uchida dates back to the summer of 1931. Accompanied by his charming wife, he paid us a visit at Iowa City, on the return trip to Japan from Berlin and Miinchen. Two years of study with Richard Goldschmidt and Karl von Frisch had made him a member of the scien tific family of Richard Hertwig .... not only by contact. Well prepared through 'years of training at the great school of Zoologists in T~kyo, he had nevertheless preserved a youthful adaptability, that enabled him to gain full advantage from his study period abroad. Iowa was suffering under a spell of sweltering heat; but even while he was wiping the sweat from his brows, our visitor denied that the weather was unbearable. Indefatigably he was carrying on the conversations .... on the lawn in the shade of the trees and in the steaming laboratories. Then and there I realized that Doctor Uchida was not only a well informed student in the widely ramified field of sex research but an investigator of originality, filled with a great enthusiasm for scientific progress. After the return to his country Professor Uchida developed his own school at the progressive Hokkaido University. It soon became known also outside of Japan, because of the wide scope and variety of subject interests as well as the quality of its scientific production. -

Chondrogenesis, Studied with the Electron Microscope
CHONDROGENESIS, STUDIED WITH THE ELECTRON MICROSCOPE GABRIEL C. GODMAN, M.1)., and KEITH R. PORTER, Ph.I). From The Rockefeller Institute. Dr. Godman's present address is Department of Microbiology, Columbia University, New York ABSTRACT The role of the cells in the fabrication of a connective tissue matrix, and the structural modifications which accompany cytodifferentiation have been investigated in developing epiphyseal cartilage of fetal rat by means of electron microscopy. Differentiation of the pre- chondral mesenchymal cells to chondroblasts is marked by the acquisition of an extensive endoplasmic reticulum, enlargement and concentration of the Golgi apparatus, the ap- pearance of membrane-bounded cytoplasmic inclusions, and the formation of specialized foci of increased density in the cell cortex. These modifications are related to the secretion of the cartilage matrix. The matrix of young hyaline cartilage consists of groups of rela- tively short, straight, banded collagen fibrils of 10 to 20 m# and a dense granular component embedded in an amorphous ground substance of moderate electron density. It is postu- lated that the first phase of fibrillogenesis takes place at the cell cortex in dense bands or striae within the ectoplasm subjacent to the cell membrane. These can be resolved into sheaves of "primary" fibrils of about 7 to 10 m#. They are supposedly shed (by excortica- tion) into the matrix space between the separating chondroblasts, where they may serve as "cores" of the definitive matrix fibrils. The diameter of the fibrils may subsequently increase up to threefold, presumably by incorporation of "soluble" or tropocollagen units from the ground substance. The chondroblast also discharges into the matrix the electron- dense amorphous or granular contents of vesicles derived from the Golgi apparatus, and the mixed contents of large vacuoles or blebs bounded by distinctive double membranes. -

Intermuscular Tendons Are Essential for the Development of Vertebrate Stomach. Ludovic Le Guen, Cécile Notarnicola, Pascal De Santa Barbara
Intermuscular tendons are essential for the development of vertebrate stomach. Ludovic Le Guen, Cécile Notarnicola, Pascal de Santa Barbara To cite this version: Ludovic Le Guen, Cécile Notarnicola, Pascal de Santa Barbara. Intermuscular tendons are essential for the development of vertebrate stomach.. Development (Cambridge, England), Company of Biologists, 2009, 136, pp.791-801. 10.1242/dev.029942. inserm-00359629 HAL Id: inserm-00359629 https://www.hal.inserm.fr/inserm-00359629 Submitted on 25 Nov 2009 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Intermuscular tendons are essential for the development of vertebrate stomach Le Guen Ludovic , Notarnicola Cécile , De Santa Barbara Pascal * Muscle et pathologies INSERM : ERI25 , IFR3 , Université Montpellier I : EA4202 , FR * Correspondence should be adressed to: Pascal De Santa Barbara <[email protected] > Abstract SUMMARY Gastrointestinal motility is ensured by the correct coordination of the enteric nervous system and the visceral smooth muscle cells (SMC), and defective development of SMCs result in gut malformations and intestinal obstructions. In order to identify the molecular mechanisms that control the differentiation of the visceral mesenchyme into SMCs in the vertebrate stomach, we developed microarrays to analyze the gene expression profiles of undifferentiated and differentiated avian stomachs. -

26 April 2010 TE Prepublication Page 1 Nomina Generalia General Terms
26 April 2010 TE PrePublication Page 1 Nomina generalia General terms E1.0.0.0.0.0.1 Modus reproductionis Reproductive mode E1.0.0.0.0.0.2 Reproductio sexualis Sexual reproduction E1.0.0.0.0.0.3 Viviparitas Viviparity E1.0.0.0.0.0.4 Heterogamia Heterogamy E1.0.0.0.0.0.5 Endogamia Endogamy E1.0.0.0.0.0.6 Sequentia reproductionis Reproductive sequence E1.0.0.0.0.0.7 Ovulatio Ovulation E1.0.0.0.0.0.8 Erectio Erection E1.0.0.0.0.0.9 Coitus Coitus; Sexual intercourse E1.0.0.0.0.0.10 Ejaculatio1 Ejaculation E1.0.0.0.0.0.11 Emissio Emission E1.0.0.0.0.0.12 Ejaculatio vera Ejaculation proper E1.0.0.0.0.0.13 Semen Semen; Ejaculate E1.0.0.0.0.0.14 Inseminatio Insemination E1.0.0.0.0.0.15 Fertilisatio Fertilization E1.0.0.0.0.0.16 Fecundatio Fecundation; Impregnation E1.0.0.0.0.0.17 Superfecundatio Superfecundation E1.0.0.0.0.0.18 Superimpregnatio Superimpregnation E1.0.0.0.0.0.19 Superfetatio Superfetation E1.0.0.0.0.0.20 Ontogenesis Ontogeny E1.0.0.0.0.0.21 Ontogenesis praenatalis Prenatal ontogeny E1.0.0.0.0.0.22 Tempus praenatale; Tempus gestationis Prenatal period; Gestation period E1.0.0.0.0.0.23 Vita praenatalis Prenatal life E1.0.0.0.0.0.24 Vita intrauterina Intra-uterine life E1.0.0.0.0.0.25 Embryogenesis2 Embryogenesis; Embryogeny E1.0.0.0.0.0.26 Fetogenesis3 Fetogenesis E1.0.0.0.0.0.27 Tempus natale Birth period E1.0.0.0.0.0.28 Ontogenesis postnatalis Postnatal ontogeny E1.0.0.0.0.0.29 Vita postnatalis Postnatal life E1.0.1.0.0.0.1 Mensurae embryonicae et fetales4 Embryonic and fetal measurements E1.0.1.0.0.0.2 Aetas a fecundatione5 Fertilization