High-Resolution Mass Spectrometry-Based Approaches for the Detection and Quantification of Peptidase Activity in Plasma
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 Evidence for Gliadin Antibodies As Causative Agents in Schizophrenia
1 Evidence for gliadin antibodies as causative agents in schizophrenia. C.J.Carter PolygenicPathways, 20 Upper Maze Hill, Saint-Leonard’s on Sea, East Sussex, TN37 0LG [email protected] Tel: 0044 (0)1424 422201 I have no fax Abstract Antibodies to gliadin, a component of gluten, have frequently been reported in schizophrenia patients, and in some cases remission has been noted following the instigation of a gluten free diet. Gliadin is a highly immunogenic protein, and B cell epitopes along its entire immunogenic length are homologous to the products of numerous proteins relevant to schizophrenia (p = 0.012 to 3e-25). These include members of the DISC1 interactome, of glutamate, dopamine and neuregulin signalling networks, and of pathways involved in plasticity, dendritic growth or myelination. Antibodies to gliadin are likely to cross react with these key proteins, as has already been observed with synapsin 1 and calreticulin. Gliadin may thus be a causative agent in schizophrenia, under certain genetic and immunological conditions, producing its effects via antibody mediated knockdown of multiple proteins relevant to the disease process. Because of such homology, an autoimmune response may be sustained by the human antigens that resemble gliadin itself, a scenario supported by many reports of immune activation both in the brain and in lymphocytes in schizophrenia. Gluten free diets and removal of such antibodies may be of therapeutic benefit in certain cases of schizophrenia. 2 Introduction A number of studies from China, Norway, and the USA have reported the presence of gliadin antibodies in schizophrenia 1-5. Gliadin is a component of gluten, intolerance to which is implicated in coeliac disease 6. -

Molecular Markers of Serine Protease Evolution
The EMBO Journal Vol. 20 No. 12 pp. 3036±3045, 2001 Molecular markers of serine protease evolution Maxwell M.Krem and Enrico Di Cera1 ment and specialization of the catalytic architecture should correspond to signi®cant evolutionary transitions in the Department of Biochemistry and Molecular Biophysics, Washington University School of Medicine, Box 8231, St Louis, history of protease clans. Evolutionary markers encoun- MO 63110-1093, USA tered in the sequences contributing to the catalytic apparatus would thus give an account of the history of 1Corresponding author e-mail: [email protected] an enzyme family or clan and provide for comparative analysis with other families and clans. Therefore, the use The evolutionary history of serine proteases can be of sequence markers associated with active site structure accounted for by highly conserved amino acids that generates a model for protease evolution with broad form crucial structural and chemical elements of applicability and potential for extension to other classes of the catalytic apparatus. These residues display non- enzymes. random dichotomies in either amino acid choice or The ®rst report of a sequence marker associated with serine codon usage and serve as discrete markers for active site chemistry was the observation that both AGY tracking changes in the active site environment and and TCN codons were used to encode active site serines in supporting structures. These markers categorize a variety of enzyme families (Brenner, 1988). Since serine proteases of the chymotrypsin-like, subtilisin- AGY®TCN interconversion is an uncommon event, it like and a/b-hydrolase fold clans according to phylo- was reasoned that enzymes within the same family genetic lineages, and indicate the relative ages and utilizing different active site codons belonged to different order of appearance of those lineages. -

Carboxypeptidase N (Kininase I) (Kdnins/Anaphylatoxins/Kallikrein/Proteases/Carboxypeptidase B) YEHUDA LEVIN*T, RANDAL A
Proc. NatL Acad. Sci. USA Vol. 79, pp. 4618-4622, August 1982 Biochemistry Isolation and characterization of the subunits of human plasma carboxypeptidase N (kininase I) (kdnins/anaphylatoxins/kallikrein/proteases/carboxypeptidase B) YEHUDA LEVIN*t, RANDAL A. SKIDGEL*, AND ERVIN G. ERDOS*t§ Departments of *Pharmacology and UInternal Medicine, University of Texas Health Science Center, 5323 Harry Hines Boulevard, Dallas, Texas 75235 Communicated by P. Kusch, May 13, 1982 ABSTRACT Carboxypeptidase N (kininase I, arginine car- MATERIALS AND METHODS boxypeptidase; EC 3.4.17.3) cleaves COOH-terminal basic amino the Parkland Me- acids of kinins, anaphylatoxins, and other peptides. The tetra- Outdated human plasma was obtained from meric enzyme of Mr 280,000 was purified from.human plasma by morial Hospital blood bank (Dallas, TX). Hippurylargininic acid ion-exchange and arginine-Sepharose affinity chromatography. (Vega Biochemicals, Tucson, AZ), bradykinin (Bachem Fine Treatment with 3 M guanidine dissociated the enzyme into sub- Chemicals, Torrance, CA), guanidine'HCI (Chemalog, South units of 83,000 and 48,000 molecular weight, which were sepa- Plainfield, NJ), trypsin (Worthington, Freehold, NJ), and hu- rated and purified by gel filtration or affinity chromatography. man plasmin [Committee on Thrombolytic Agents (CTA) Stan- When tested with hippurylarginine, hippurylargininic acid, ben- dard, 10 units/ml, the American National'Red Cross] were used zoylalanyllysine, or bradyldkini the Mr 48,000 subunit was as ac- as received. Human urinary kallikrein was donated by H. Fritz tive as the intact enzyme-and was easily distinguished from human (Munich, Federal Republic of Germany) and human plasma pancreatic carboxypeptidase B (EC 3.4.17.2). -

JASON MARC GOLDSTEIN the Isolation, Characterization
JASON MARC GOLDSTEIN The Isolation, Characterization and Cloning of Three Novel Peptidases From Streptoccocus gordonii: Their Potential Roles in Subacute Bacterial Endocarditis (Under the Direction of JAMES TRAVIS) Streptococcus gordonii is generally considered a benign inhabitant of the oral microflora yet is a primary etiological agent in the development of subacute bacterial endocarditis (SBE), an inflammatory state that propagates thrombus formation and tissue damage on the surface of heart valves. Colonization and adherence mechanisms have been identified, yet factors necessary to sustain growth remain unidentified. Strain FSS2 produced three extracellular aminopeptidase activities during growth in neutral pH- controlled batch cultures. The first included a serine-class dipeptidyl-aminopeptidase, an x-Pro DPP (Sg-xPDPP) found as an 85 kDa monomer by SDS-PAGE while appearing as a homodimer under native conditions. Kinetic studies indicated a unique and stringent x- Pro specificity comparable to the DPPIV/CD26 and lactococcal x-Pro DPP families. Isolation of the full-length gene uncovered a 759-amino acid polypeptide with a mass of 87,115 Da and theoretical pI of 5.6. Significant homology was found with PepX gene family members from Lactobacillus ssp. and Lactococcus ssp., and putative streptococcal x-Pro DPPs. The second activity was a putative serine-class arginine aminopeptidase (Sg- RAP) with some cysteine-class characteristics. It was found as a protein monomer of 70 kDa under denaturing conditions. Nested PCR cloning enabled the isolation of a 324 bp- long DNA fragment encoding the protein’s 108 amino acid N-terminus. Culture activity profiles and N-terminal sequence analysis indicated the release of this protein from the cell surface. -

Table 6. Putative Genes Involved in the Utilization of Carbohydrates in G
Table 6. Putative genes involved in the utilization of carbohydrates in G. thermodenitrificans NG80-2 genome Carbohydrates* Enzymes Gene ID Glycerol Glycerol Kinase GT1216 Glycerol-3-phosphate dehydrogenase, aerobic GT2089 NAD(P)H-dependent glycerol-3-phosphate dehydrogenase GT2153 Enolase GT3003 2,3-bisphosphoglycerate-independentphosphoglycerate mutase GT3004 Triosephosphate isomerase GT3005 3-phosphoglycerate kinase GT3006 Glyceraldehyde-3-phosphate dehydrogenase GT3007 Phosphoglycerate mutase GT1326 Pyruvate kinase GT2663 L-Arabinose L-arabinose isomerase GT1795 L-ribulokinase GT1796 L-ribulose 5-phosphate 4-epimerase GT1797 D-Ribose Ribokinase GT3174 Transketolase GT1187 Ribose 5-phosphate epimerase GT3316 D-Xylose Xylose kinase GT1756 Xylose isomerase GT1757 D-Galactose Galactokinase GT2086 Galactose-1-phosphate uridyltransferase GT2084 UDP-glucose 4-epimerase GT2085 Carbohydrates* Enzymes Gene ID D-Fructose 1-phosphofructokinase GT1727 Fructose-1,6-bisphosphate aldolase GT1805 Fructose-1,6-bisphosphate aldolase type II GT3331 Triosephosphate isomerase GT3005 D-Mannose Mannnose-6 phospate isomelase GT3398 6-phospho-1-fructokinase GT2664 D-Mannitol Mannitol-1-phosphate dehydrogenase GT1844 N-Acetylglucosamine N-acetylglucosamine-6-phosphate deacetylase GT2205 N-acetylglucosamine-6-phosphate isomerase GT2204 D-Maltose Alpha-1,4-glucosidase GT0528, GT1643 Sucrose Sucrose phosphorylase GT3215 D-Trehalose Alpha-glucosidase GT1643 Glucose kinase GT2381 Inositol Myo-inositol catabolism protein iolC;5-dehydro-2- GT1807 deoxygluconokinase -

An Investigation on Catalysis of Acylaminoacyl Peptidases
An investigation on catalysis of acylaminoacyl peptidases PhD Thesis András László Kiss Doctorate School of Biology School leader: Prof. Anna Erdei Structural Biochemistry Programme Programme leader: Prof. László Gráf Supervisor: Prof. László Polgár Institute of Enzymology, Biological Research Center, Hungarian Academy of Sciences Budapest 2007 1. Introduction Serine peptidases contain two residues at the active site in addition to the catalytic triad (His, Asp, Ser), which form a cavity called “oxyanion hole” that accommodates the negatively charged oxyanion in the transition state of the catalysis and donate two H- bonds. Enzymes of the prolyl oligopeptidase (POP) family (acylaminoacyl peptidase, prolyl oligopeptidase, dipeptidyl peptidase IV and oligopeptidase B) are extensively studied. These enzymes are larger than classical serine peptidases and are composed of a peptidase domain with an α/β hydrolase fold and a β-propeller domain. POP and oligopeptidase B are monomeric enzymes and endopeptidases, while the exopeptidase acylaminoacyl peptidase and dipeptidyl peptidase IV are tetrameric and dimeric enzymes, respectively. Acylaminoacyl peptidase (AAP) cleaves acylated amino acids from the N-terminus of the N-acylated peptides that plays important role in many biological and disease processes. Human AAP is encoded by the DNF15S2 locus on the short arm of chromosome 3 at the region 21, which suffers deletions in small cell lung carcinomas, and renal carcinomas, resulting in deficiency in the expression of the enzyme. Acylaminoacyl peptidase is also supposed to be involved in the degradation of oxidatively damaged proteins in cells and can be associated with various diseases where damaged proteins aggregate. Its involvement in cataract formation and in the breakdown of immunogenic formylmethionyl-peptides in the digestive tract after bacterial attack was also suggested. -

Structure and Function of a Serine Carboxypeptidase Adapted for Degradation of the Protein Synthesis Antibiotic Microcin C7
Structure and function of a serine carboxypeptidase adapted for degradation of the protein synthesis antibiotic microcin C7 Vinayak Agarwala,b, Anton Tikhonovc,d, Anastasia Metlitskayac, Konstantin Severinovc,d,e, and Satish K. Naira,b,f,1 aCenter for Biophysics and Computational Biology, bInstitute for Genomic Biology, and fDepartment of Biochemistry, University of Illinois at Urbana-Champaign, 600 South Mathews Avenue, Urbana, IL 61801; cInstitutes of Molecular Genetics and Gene Biology, Russian Academy of Sciences, Moscow 11934, Russia; eDepartment of Molecular Biology and Biochemistry, and dWaksman Institute, Rutgers, State University of New Jersey, Piscataway, NJ 08854 Edited by Perry Allen Frey, University of Wisconsin, Madison, WI, and approved January 20, 2012 (received for review August 30, 2011) Several classes of naturally occurring antimicrobials exert their activity is exerted after intracellular processing. Examples of antibiotic activity by specifically targeting aminoacyl-tRNA synthe- naturally occurring antibiotics that employ this Trojan horse strat- tases, validating these enzymes as drug targets. The aspartyl tRNA egy include the LeuRS inhibitor agrocin 84 (in which the toxic synthetase “Trojan horse” inhibitor microcin C7 (McC7) consists of a group is linked through a phosphoramidate bond to a D-glucofur- nonhydrolyzable aspartyl-adenylate conjugated to a hexapeptide anosyloxyphosphoryl moiety) (5), the SerRS inhibitor albomycin carrier that facilitates active import into bacterial cells through an (a toxic group covalently linked to a hydroxymate siderophore) oligopeptide transport system. Subsequent proteolytic processing (6), and the AspRS inhibitor microcin C7 (Fig. 1A, 1) (McC7; releases the toxic compound inside the cell. Producing strains consisting of a modified aspartyl-adenylate linked to a six-residue of McC7 must protect themselves against autotoxicity that may re- peptide carrier). -

Two Distinct Gene Subfamilies Within the Family of Cysteine Protease Genes (Tetrahymena/Propeptide/Cathepsin) KATHLEEN M
Proc. Natl. Acad. Sci. USA Vol. 90, pp. 3063-3067, April 1993 Biochemistry Two distinct gene subfamilies within the family of cysteine protease genes (tetrahymena/propeptide/cathepsin) KATHLEEN M. KARRER*, STACIA L. PEIFFERt, AND MICHELE E. DITOMAS Department of Biology, Marquette University, Milwaukee, WI 53233 Communicated by David M. Prescott, January 7, 1993 ABSTRACT A cDNA clone for a physiologically regulated (4, 5). The clone was isolated from a cDNA library of RNA Tetrahymena cysteine protease gene was sequenced. The nu- from starved cells cloned into the Pst I site ofpUC9 (4). DNA cleotide sequence predicts that the clone encodes a 336-amino fragments were subcloned into pBluescript for sequencing. acid protein composed of a 19-residue N-terminal signal se- The sequence was scanned for open reading frames by using quence followed by a 107-residue propeptide and a 210-residue the DNA INSPECTOR IIE program (Textco), taking into con- mature protein. Comparison of the deduced amino acid se- sideration that in Tetrahymena, as in several ciliates, TAA quence of the protein with those of other cysteine proteases and TAG code for Gln (6-8). DNA sequences that code for revealed a highly conserved interspersed amino acid motif in homologous proteins were identified through a Pearson and the propeptide region of the protein, the ERFNIN motif. The Lipman (9) search ofthe EMBL/GenBank data base by using motifwas present in all ofthe cysteine proteases in the data base with the exception of the cathepsin B-like proteins, which have the TFASTA program. shorter propeptides. Differences in the propeptides and in conserved amino acids of the mature proteins suggest that the RESULTS ERFNIN proteases and the cathepsin B-like proteases consti- tute two distinct subfamilies within the cysteine proteases. -
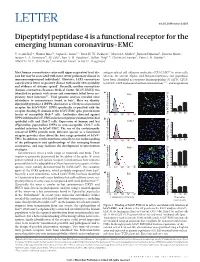
Dipeptidyl Peptidase 4 Is a Functional Receptor for the Emerging Human Coronavirus-EMC
LETTER doi:10.1038/nature12005 Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC V. Stalin Raj1*, Huihui Mou2*, Saskia L. Smits1,3, Dick H. W. Dekkers4, Marcel A. Mu¨ller5, Ronald Dijkman6, Doreen Muth5, Jeroen A. A. Demmers4, Ali Zaki7, Ron A. M. Fouchier1, Volker Thiel6,8, Christian Drosten5, Peter J. M. Rottier2, Albert D. M. E. Osterhaus1, Berend Jan Bosch2 & Bart L. Haagmans1 Most human coronaviruses cause mild upper respiratory tract dis- antigen-related cell adhesion molecules (CEACAM)10 to enter cells, ease but may be associated with more severe pulmonary disease in whereas for several Alpha- and Betacoronaviruses, two peptidases immunocompromised individuals1. However, SARS coronavirus have been identified as receptors (aminopeptidase N (APN, CD13) caused severe lower respiratory disease with nearly 10% mortality for hCoV-229E and several animal coronaviruses11,12, and angiotensin and evidence of systemic spread2. Recently, another coronavirus (human coronavirus-Erasmus Medical Center (hCoV-EMC)) was a identified in patients with severe and sometimes lethal lower res- )] Vero –1 piratory tract infection3,4. Viral genome analysis revealed close 9 ml 8 5 50 relatedness to coronaviruses found in bats . Here we identify 7 dipeptidyl peptidase 4 (DPP4; also known as CD26) as a functional 6 5 receptor for hCoV-EMC. DPP4 specifically co-purified with the 4 receptor-binding S1 domain of the hCoV-EMC spike protein from 3 Relative cell number log[GE (TCID 0 20 40 lysates of susceptible Huh-7 cells. Antibodies directed against 0 101 102 103 104 DPP4 inhibited hCoV-EMC infection of primary human bronchial b )] COS-7 –1 epithelial cells and Huh-7 cells. -

And Exopeptidases in a Processing Enzyme System: Activation
Proc. Nail. Acad. Sci. USA Vol. 85, pp. 5468-5472, August 1988 Biochemistry Relationship between endo- and exopeptidases in a processing enzyme system: Activation of an endoprotease by the aminopeptidase B-like activity in somatostatin-28 convertase (brain cortex/basic amino acid pairs/peptide substrates/protease inhibitors/prohormone maturation) SOPHIE GOMEZ, PABLO GLUSCHANKOF, AGNES LEPAGE, AND PAUL COHEN Groupe de Neurobiochimie Cellulaire et Moldculaire, Universitd Pierre et Marie Curie, Unitd Associde 554 au Centre National de la Recherche Scientifique, 96 boulevard Raspail, 75006 Paris, France Communicated by I. Robert Lehman, April 8, 1988 (receivedfor review December 15, 1987) ABSTRACT The somatostatin-28 convertase activity in- somatostatin-14 and the amino-terminal dodecapeptide so- volved in vitro in the processing of somatostatin-28 into the matostatin-28-(1-12) (16). neuropeptides somatostatin-28-(1-12) and somatostatin-14 is We have described an endoprotease that cleaves the composed of an endoprotease and a basic aminopeptidase. We peptide bond on the amino side ofthe Arg-Lys doublet in the report herein on the purification to apparent homogeneity of somatostatin-28 sequence (17, 18), releasing the somato- these two constituents and on their functional interrelationship. statin-28-(1-12) fragment and [Arg-2,Lys-1]somatostatin-14. In particular we observed that after various physicochemical The released [Arg-2,Lys-']somatostatin-14 is further proc- treatments, the 90-kDa endoprotease activity was recovered essed by an aminopeptidase B-like activity (18, 19) that is also both at this molecular mass and as a 45-kDa entity. Moreover, present in the preparation. We report herein the purification the production of [Arg2,LysJllsomatostatin-14 from somato- to apparent homogeneity ofthese two activities. -

Proteases for Biocatalysis
Proteases for biocatalysis for smarter chemical synthesis Biocatalysis Biocatalysis involves the implementation of natural catalysts, such as enzymes, in place of chemical catalysts in synthetic processes. Compared to chemical catalysts, enzymes offer: • higher reaction rates • milder reaction conditions • high reaction specificity with no side products This change can enable new, more sustainable routes for the production of intermediates and active pharmaceutical ingredients (APIs). However, please note Novozymes products do not comply with manufacturing according to pharmaceutical standards and Novozymes products must not be used as active pharmaceutical ingredients (APIs) or excipients. Biocatalysis has become an increasingly important tool for medicinal, process and polymer chemists, allowing the development of efficient and highly attractive synthetic processes on an industrial scale. Use of enzymes in catalysis is a well-established technology within the chemical industry. An advantage of enzymes in organic synthesis is their remarkable selective properties, which provide commercial benefits including: • high selectivity in production of single stereoisomers • fewer side reactions • less reprocessing and purification steps • easier product separation • less pollution The combination of all of these advantages leads to a reduction in costs. Enzyme catalysts work by lowering the activation energy (Ea‡) for a reaction, thus dramatically increasing the rate of the reaction. As a result, products are formed faster and reactions reach their equilibrium state more rapidly. Most enzyme reaction rates are millions of times faster than those of comparable uncatalyzed reactions. As with all catalysts, enzymes are not consumed by the reactions they catalyze, nor do they alter the equilibrium of these reactions. However, enzymes do differ from most other catalysts in that they are highly specific for their substrates. -

Chapter 11 Cysteine Proteases
CHAPTER 11 CYSTEINE PROTEASES ZBIGNIEW GRZONKA, FRANCISZEK KASPRZYKOWSKI AND WIESŁAW WICZK∗ Faculty of Chemistry, University of Gdansk,´ Poland ∗[email protected] 1. INTRODUCTION Cysteine proteases (CPs) are present in all living organisms. More than twenty families of cysteine proteases have been described (Barrett, 1994) many of which (e.g. papain, bromelain, ficain , animal cathepsins) are of industrial impor- tance. Recently, cysteine proteases, in particular lysosomal cathepsins, have attracted the interest of the pharmaceutical industry (Leung-Toung et al., 2002). Cathepsins are promising drug targets for many diseases such as osteoporosis, rheumatoid arthritis, arteriosclerosis, cancer, and inflammatory and autoimmune diseases. Caspases, another group of CPs, are important elements of the apoptotic machinery that regulates programmed cell death (Denault and Salvesen, 2002). Comprehensive information on CPs can be found in many excellent books and reviews (Barrett et al., 1998; Bordusa, 2002; Drauz and Waldmann, 2002; Lecaille et al., 2002; McGrath, 1999; Otto and Schirmeister, 1997). 2. STRUCTURE AND FUNCTION 2.1. Classification and Evolution Cysteine proteases (EC.3.4.22) are proteins of molecular mass about 21-30 kDa. They catalyse the hydrolysis of peptide, amide, ester, thiol ester and thiono ester bonds. The CP family can be subdivided into exopeptidases (e.g. cathepsin X, carboxypeptidase B) and endopeptidases (papain, bromelain, ficain, cathepsins). Exopeptidases cleave the peptide bond proximal to the amino or carboxy termini of the substrate, whereas endopeptidases cleave peptide bonds distant from the N- or C-termini. Cysteine proteases are divided into five clans: CA (papain-like enzymes), 181 J. Polaina and A.P. MacCabe (eds.), Industrial Enzymes, 181–195.