Sorbaria Species Cultivated in Poland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Phylogeny of Maleae (Rosaceae) Based on Multiple Chloroplast Regions: Implications to Genera Circumscription
Hindawi BioMed Research International Volume 2018, Article ID 7627191, 10 pages https://doi.org/10.1155/2018/7627191 Research Article Phylogeny of Maleae (Rosaceae) Based on Multiple Chloroplast Regions: Implications to Genera Circumscription Jiahui Sun ,1,2 Shuo Shi ,1,2,3 Jinlu Li,1,4 Jing Yu,1 Ling Wang,4 Xueying Yang,5 Ling Guo ,6 and Shiliang Zhou 1,2 1 State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of the Chinese Academy of Sciences, Beijing 100043, China 3College of Life Science, Hebei Normal University, Shijiazhuang 050024, China 4Te Department of Landscape Architecture, Northeast Forestry University, Harbin 150040, China 5Key Laboratory of Forensic Genetics, Institute of Forensic Science, Ministry of Public Security, Beijing 100038, China 6Beijing Botanical Garden, Beijing 100093, China Correspondence should be addressed to Ling Guo; [email protected] and Shiliang Zhou; [email protected] Received 21 September 2017; Revised 11 December 2017; Accepted 2 January 2018; Published 19 March 2018 Academic Editor: Fengjie Sun Copyright © 2018 Jiahui Sun et al. Tis is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Maleae consists of economically and ecologically important plants. However, there are considerable disputes on generic circumscription due to the lack of a reliable phylogeny at generic level. In this study, molecular phylogeny of 35 generally accepted genera in Maleae is established using 15 chloroplast regions. Gillenia isthemostbasalcladeofMaleae,followedbyKageneckia + Lindleya, Vauquelinia, and a typical radiation clade, the core Maleae, suggesting that the proposal of four subtribes is reasonable. -

A Molecular Phylogeny of Selected Species of Genus Prunus L
African Journal of Biotechnology Vol. 9(31), pp. 4867-4872, 2 August, 2010 Available online at http://www.academicjournals.org/AJB DOI: 10.5897/AJB10.519 ISSN 1684–5315 ©2010 Academic Journals Full Length Research Paper A molecular phylogeny of selected species of genus Prunus L. (Rosaceae) from Pakistan using the internal transcribed spacer (ITS) spacer DNA Syed Aneel Gilani1*, Rizwana Aleem Qureshi1, Amir M. Khan1 and Daniel Potter2 1Department of Plant Sciences, Quaid-I-Azam University, Islamabad, Pakistan. 2Department of Plant Sciences, University of California, Davis 95616, USA. Accepted 30 June, 2010 Prunus is found in all four provinces of Pakistan, that is, Punjab, NWFP, Sindh and Baluchistan including Azad Kashmir region. Studies on the family Rosaceae is scanty in the Flora of Pakistan and there is a lot of taxonomic work yet to be done, for the proper classification and placement of different genera under different sub-families. In the present study, the genus Prunus was studied in detail to find out the phylogenetic relationship among the 23 species of Prunus, selected from different regions of Pakistan and GenBank using maximum parsimony analysis of sequence polymorphism in nuclear ITS-9 and ITS-6 spacer DNA. The results for the internal transcribed spacer (ITS)- 9 and ITS- 6 primers confirm the work done by early phylogenetists with additions of new species from Pakistan including Prunus bokhariensis, Prunus dulcis (Mill.) D.A. Webb. (Syn. Prunus amygdalus) and Prunus cornuta (Wall. ex. Royle) Steudel. These are indigenous to Pakistan. In the ITS strict consensus results for example, the clade consisting of Laurocerasus, Padus and Cerasus subgenera are sister to the rest of the clades in the phylogenetic tree. -

Phylogenetic Relationships in Rosaceae Inferred from Chloroplast Matk and Trnl-Trnf Nucleotide Sequence Data
Plant Syst. Evol. 231: 77±89 32002) Phylogenetic relationships in Rosaceae inferred from chloroplast matK and trnL-trnF nucleotide sequence data D. Potter1, F. Gao1, P. Esteban Bortiri1, S.-H. Oh1, and S. Baggett2 1Department of Pomology, University of California, Davis, USA 2Ph.D. Program Biology, Lehman College, City University of New York, New York, USA Received February 27, 2001 Accepted October 11, 2001 Abstract. Phylogenetic relationships in Rosaceae economically important fruits of temperate were studied using parsimony analysis of nucleo- regions is produced by members of this family, tide sequence data from two regions of the including species of Malus 3apples), Pyrus chloroplast genome, the matK gene and the trnL- 3pears), Prunus 3plums, peaches, cherries, trnF region. As in a previously published phylog- almonds, and apricots), Rubus 3raspberries), eny of Rosaceae based upon rbcL sequences, and Fragaria 3strawberries). The family also monophyletic groups were resolved that corre- includes many ornamentals, e.g., species of spond, with some modi®cations, to subfamilies Maloideae and Rosoideae, but Spiraeoideae were Rosa 3roses), Potentilla 3cinquefoil), and polyphyletic. Three main lineages appear to have Sorbus 3mountain ash). A variety of growth diverged early in the evolution of the family: 1) habits, fruit types, and chromosome numbers Rosoideae sensu stricto, including taxa with a base is found within the family 3Robertson 1974), chromosome number of 7 3occasionally 8); 2) which is traditionally divided into four sub- actinorhizal Rosaceae, a group of taxa that engage families on the basis of fruit type 3e.g., Schulze- in symbiotic nitrogen ®xation; and 3) the rest of the Menz 1964). -

The Emergence of Core Eudicots: New Floral Evidence from the Earliest
Downloaded from http://rspb.royalsocietypublishing.org/ on January 18, 2017 The emergence of core eudicots: rspb.royalsocietypublishing.org new floral evidence from the earliest Late Cretaceous Else Marie Friis1, Kaj Raunsgaard Pedersen2 and Peter R. Crane3,4 Research 1Department of Palaeobiology, Swedish Museum of Natural History, Stockholm, Sweden 2 Cite this article: Friis EM, Pedersen KR, Crane Department of Geoscience, University of Aarhus, Aarhus, Denmark 3Yale School of Forestry and Environmental Studies, New Haven, CT, USA PR. 2016 The emergence of core eudicots: 4Oak Spring Garden Foundation, Upperville, VA, USA new floral evidence from the earliest EMF, 0000-0003-2936-2761 Late Cretaceous. Proc. R. Soc. B 283: 20161325. http://dx.doi.org/10.1098/rspb.2016.1325 Eudicots, the most diverse of the three major clades of living angiosperms, are first recognized in the latest Barremian–earliest Aptian. All Early Cretaceous forms appear to be related to species-poor lineages that diverged before the rise of core eudicots, which today comprise more than 70% of angiosperm Received: 11 June 2016 species. Here, we report the discovery of a well-preserved flower, Caliciflora Accepted: 14 November 2016 mauldinensis, from the earliest Late Cretaceous, with unequivocal core eudicot features, including five sepals, five petals and two whorls of stamens borne on the rim of a floral cup containing three free carpels. Pollen is tricolporate. Carpels mature into follicular fruitlets. This character combination suggests a phylogenetic position among rosids, but more specific assignment is Subject Areas: precluded by complex patterns of character evolution among the very large evolution, palaeontology, structural biology number of potentially relevant extant taxa. -

New York Non-Native Plant Invasiveness Ranking Form
NEW YORK NON-NATIVE PLANT INVASIVENESS RANKING FORM Scientific name: Sorbaria sorbifolia (L.) A. Braun USDA Plants Code: SOSO2 Common names: False spiraea Native distribution: Eastern and northern Asia Date assessed: December 1, 2010 Assessors: Gerry Moore Reviewers: LIISMA SRC Date Approved: 10 January 2011 Form version date: 10 July 2009 New York Invasiveness Rank: Unknown (fewer than 70 total points assessed) Distribution and Invasiveness Rank (Obtain from PRISM invasiveness ranking form) PRISM Status of this species in each PRISM: Current Distribution Invasiveness Rank 1 Adirondack Park Invasive Program Not Assessed Not Assessed 2 Capital/Mohawk Not Assessed Not Assessed 3 Catskill Regional Invasive Species Partnership Not Assessed Not Assessed 4 Finger Lakes Not Assessed Not Assessed 5 Long Island Invasive Species Management Area Common Unknown 6 Lower Hudson Not Assessed Not Assessed 7 Saint Lawrence/Eastern Lake Ontario Not Assessed Not Assessed 8 Western New York Not Assessed Not Assessed Invasiveness Ranking Summary Total (Total Answered*) Total (see details under appropriate sub-section) Possible 1 Ecological impact 40 (0) 0 2 Biological characteristic and dispersal ability 25 (22) 17 3 Ecological amplitude and distribution 25 (21) 19 4 Difficulty of control 10 (10) 8 b a Outcome score 100 (53) 44 † Relative maximum score § New York Invasiveness Rank Unknown (fewer than 70.00 total points assessed) * For questions answered “unknown” do not include point value in “Total Answered Points Possible.” If “Total Answered Points Possible” is less than 70.00 points, then the overall invasive rank should be listed as “Unknown.” †Calculated as 100(a/b) to two decimal places. -
Rosaceous Chamaebatiaria-Like Foliage from the Paleogene of Western North America Jack A
Aliso: A Journal of Systematic and Evolutionary Botany Volume 12 | Issue 1 Article 14 1988 Rosaceous Chamaebatiaria-Like Foliage from the Paleogene of Western North America Jack A. Wolfe U.S. Geological Survey Wesley Wehr University of Washington Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Botany Commons Recommended Citation Wolfe, Jack A. and Wehr, Wesley (1988) "Rosaceous Chamaebatiaria-Like Foliage from the Paleogene of Western North America," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 12: Iss. 1, Article 14. Available at: http://scholarship.claremont.edu/aliso/vol12/iss1/14 ALISO 12(1), 1988, pp. 177-200 ROSACEOUS CHAMAEBATIARIA-LIKE FOLIAGE FROM THE PALEOGENE OF WESTERN NORTH AMERICA JACK A. WOLFE U.S. Geological Survey, MS-919, Federal Center, Denver, Colorado 80225 AND WESLEY WEHR Burke Memorial Washington State Museum University of Washington. Seattle, Washington 98195 ABSTRACT Chamaebatiaria and Chamaebatia, two characteristic genera of the Californian floristic province, are traditionally placed in different subfamilies ofRosaceae , Spiraeoideae and Rosoideae, respect ively. Analysis of the foliar and reproductive characters of the extant species of these genera indicates that the two genera could be closely related and the assignment of Chamaebatia to Rosoideae invalid. Fossil leaves oflineages ofboth genera occur in the Paleogene montane floras ofthe Rocky Mountain region and provide evidence that the two lineages diverged from a common ancestor in the Eocene. The common ancestor probably was adapted to sunny habitats in mesic coniferous forest, and , during the post-Eocene, the two lineages were able to adapt to progressively drier climates. A third extant genus, the east Asian Sorbaria, also appears to be closely related to the California genera and to have been derived from the same common ancestor. -

Diversity and Evolution of Rosids
Diversity and Evolution of Rosids . roses, currants, peonies . Eudicots • continue survey through the eudicots or tricolpates • vast majority of eudicots are Rosids (polypetalous) and Asterids core (sympetalous) eudicots rosid asterid Eudicots • unlike Asterids, Rosids (in orange) now represent a diverse set of families *Saxifragales • before examining the large Rosid group, look at a small but important order of flowering plants - Saxifragales Paeonia Sedum *Saxifragales • small group of 16 families and about 2500 species sister to Rosids • ancient lineage from 120 mya and underwent rapid radiation Paeonia Sedum *Saxifragales • part of this ancient radiation may involve this small family of holo-parasites - Cynomoriaceae *Saxifragales • they generally can be identified by their two or more separate or semi-fused carpels, but otherwise quite variable Paeonia Sedum Paeoniaceae 1 genus / 33 species • like many of these families, Paeonia exhibits an Arcto-Tertiary distribution Paeoniaceae 1 genus / 33 species • small shrubs with primitive features of perianth and stamens • hypogynous with 5-8 separate carpels developing into follicles Cercidiphyllaceae 1 genus / 2 species • small trees (kadsura-tree) restricted to eastern China and Japan . • . but fossils in North America and Europe from Tertiary Cercidiphyllaceae 1 genus / 2 species • unisexual, wind-pollinated but do produce follicles Hamamelidaceae 27 genera and 80 species - witch hazels • family of trees and shrubs in subtropical and temperate areas but only 1 species in Wisconsin - witch -

1. Rosaceae: Taxonomy, Economic Importance, Genomics
1. Rosaceae: Taxonomy, Economic Importance, Genomics Kim E. Hummer and Jules Janick A rose by any other name would smell as sweet. Shakespeare A rose is a rose is a rose. Gertrude Stein The Rose Family The rose is a rose And was always a rose; But the theory now goes That the apple’s a rose, And the pear is, and so’s The plum, I suppose. The dear only knows What will next prove a rose. You, of course, are a rose, But were always a rose. Robert Frost 1 Nomenclature and Taxonomy 1.1 Origins The magnificent simplicity, or to some, the monotonous consistency, of the actin- iomorphic flowers of the rose family has been recognized for millennia. The origin of the name rose is summarized in the American Heritage Dictionary (2000): The English word rose comes from Latin and Old French. Latin rosa may be an Etruscan form of Greek Rhodia, “Rhodian, originating from Rhodes.” The Attic Greek word for rose K.E. Hummer (B) U. S. Department of Agriculture, Agricultural Research Service, National Clonal Germplasm Repository, 33447 Peoria Road, Corvallis, Oregon, 97333, USA K.M. Folta, S.E. Gardiner (eds.), Genetics and Genomics of Rosaceae, Plant Genetics 1 and Genomics: Crops and Models 6, DOI 10.1007/978-0-387-77491-6 1, C Springer Science+Business Media, LLC 2009 2 K.E. Hummer and J. Janick is rhodon, and in Sappho’s Aeolic dialect of Greek it is wrodon. In Avestan, the language of the Persian prophet Zoroaster, “rose” is varda and in Armenian vard, words both related to the Aeolic form. -

S Genera Layout 1/31/08 12:45 PM Page 997
S genera Layout 1/31/08 12:45 PM Page 997 Arecaceae—Palm family S Sabal Adans. palmetto David F. Olson, Jr., R. L. Barnes, and Robert P.Karrfalt Drs. Olson and Barnes retired from the USDA Forest Service’s Southeastern Forest Experiment Station; Mr. Karrfalt is director of the USDA Forest Service’s National Seed Laboratory, Dry Branch, Georgia Growth habits, occurrence, and use. Palmettos— of about 1.3 m (Bailey 1939; McCurrach 1960). It has a genus Sabal—are native to the Western Hemisphere and are restricted range in the dry pinelands and scrub of central distributed from the Bermuda Islands and the South Atlantic Florida (Small 1933). The bud is eaten as a salad vegetable, and Gulf States through the West Indies to Venezuela and and the fruits are eaten by animals and birds. Mexico (Sargent 1965). Five species inhabit the southeastern Flowering and fruiting. The perfect white flowers of United States, Puerto Rico, and the Virgin Islands (table 1). cabbage palmetto measure about 6 mm in diameter and are Cabbage palmetto has tree form and attains a height at matu- borne in drooping clusters 1.3 to 1.8 m long from June to rity of 12 to 27 m (Sargent 1965); it is found from North August, depending upon latitude (Sargent 1965; Snyder Carolina to south Florida, in low flatwoods and on offshore 1952; West and Arnold 1947). The flowers are pollinated by islands in the north, and becoming common throughout the insects (Knuth 1906). The fruit is a berry, subglobose or lower part of the Florida peninsula. -
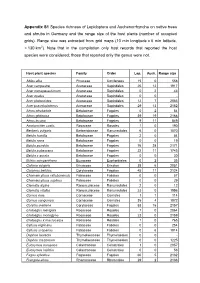
Appendix S1 Species Richness of Lepidoptera and Auchenorrhyncha on Native Trees and Shrubs in Germany and the Range Size of the Host Plants (Number of Occupied Grids)
Appendix S1 Species richness of Lepidoptera and Auchenorrhyncha on native trees and shrubs in Germany and the range size of the host plants (number of occupied grids). Range size was extracted from grid maps (10 min longitude x 6 min latitude, ≈ 130 km 2). Note that in the compilation only host records that reported the host species were considered; those that reported only the genus were not. Host plant species Family Order Lep. Auch. Range size Abies alba Pinaceae Coniferales 15 0 558 Acer campestre Aceraceae Sapindales 25 12 1917 Acer monspessulanum Aceraceae Sapindales 0 2 43 Acer opalus Aceraceae Sapindales 0 0 - Acer platanoides Aceraceae Sapindales 12 7 2083 Acer pseudoplatanus Aceraceae Sapindales 29 13 2152 Alnus alnobetula Betulaceae Fagales 0 2 54 Alnus glutinosa Betulaceae Fagales 39 19 2168 Alnus incana Betulaceae Fagales 9 11 849 Amelanchier ovalis Rosaceae Rosales 1 0 190 Berberis vulgaris Berberidaceae Ranunculales 6 0 1070 Betula humilis Betulaceae Fagales 2 0 54 Betula nana Betulaceae Fagales 0 0 19 Betula pendula Betulaceae Fagales 76 28 2171 Betula pubescens Betulaceae Fagales 22 11 1745 Betula x aurata Betulaceae Fagales 0 0 30 Buxus sempervirens Buxaceae Euphorbiales 0 2 35 Calluna vulgaris Ericaceae Ericales 38 6 2051 Carpinus betulus Corylaceae Fagales 45 11 2124 Chamaecytisus ratisbonensis Fabaceae Fabales 0 0 57 Chamaecytisus supinus Fabaceae Fabales 0 0 29 Clematis alpina Ranunculaceae Ranunculales 2 0 12 Clematis vitalba Ranunculaceae Ranunculales 23 0 1556 Cornus mas Cornaceae Cornales 1 1 114 Cornus sanguinea -

The Emergence of Core Eudicots: New Floral Evidence from the Earliest
The emergence of core eudicots: rspb.royalsocietypublishing.org new floral evidence from the earliest Late Cretaceous Else Marie Friis1, Kaj Raunsgaard Pedersen2 and Peter R. Crane3,4 Research 1Department of Palaeobiology, Swedish Museum of Natural History, Stockholm, Sweden 2 Cite this article: Friis EM, Pedersen KR, Crane Department of Geoscience, University of Aarhus, Aarhus, Denmark 3Yale School of Forestry and Environmental Studies, New Haven, CT, USA PR. 2016 The emergence of core eudicots: 4Oak Spring Garden Foundation, Upperville, VA, USA new floral evidence from the earliest EMF, 0000-0003-2936-2761 Late Cretaceous. Proc. R. Soc. B 283: 20161325. http://dx.doi.org/10.1098/rspb.2016.1325 Eudicots, the most diverse of the three major clades of living angiosperms, are first recognized in the latest Barremian–earliest Aptian. All Early Cretaceous forms appear to be related to species-poor lineages that diverged before the rise of core eudicots, which today comprise more than 70% of angiosperm Received: 11 June 2016 species. Here, we report the discovery of a well-preserved flower, Caliciflora Accepted: 14 November 2016 mauldinensis, from the earliest Late Cretaceous, with unequivocal core eudicot features, including five sepals, five petals and two whorls of stamens borne on the rim of a floral cup containing three free carpels. Pollen is tricolporate. Carpels mature into follicular fruitlets. This character combination suggests a phylogenetic position among rosids, but more specific assignment is Subject Areas: precluded by complex patterns of character evolution among the very large evolution, palaeontology, structural biology number of potentially relevant extant taxa. The whorled floral organization is consistent with ideas that this stable pattern evolved early and was a pre- Keywords: requisite for more integrated patterns of floral architecture that evolved fossil flower, rosids, asterid, SRXTM, later. -

Vascular Plant Inventory for Lewis and Clark National Historical Park Public Version
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science Vascular Plant Inventory for Lewis and Clark National Historical Park Public Version Natural Resource Technical Report NPS/LEWI/NRTR—2012/603.N ON THE COVER Headland at Cape Disappointment Photograph by: Lindsey Koepke Wise Vascular Plant Inventory for Lewis and Clark National Historical Park Public Version Natural Resource Technical Report NPS/LEWI/NRTR—2012/603.N Lindsey Koepke Wise Oregon Biodiversity Information Center Institute for Natural Resources / INR Portland State University P.O. Box 751 Portland, OR 97207 Jimmy Kagan Oregon Biodiversity Information Center Institute for Natural Resources / INR Portland State University P.O. Box 751 Portland, OR 97207 July 2012 U.S. Department of the Interior National Park Service Natural Resource Stewardship and Science Fort Collins, Colorado The National Park Service, Natural Resource Stewardship and Science office in Fort Collins, Colorado publishes a range of reports that address natural resource topics of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Technical Report Series is used to disseminate results of scientific studies in the physical, biological, and social sciences for both the advancement of science and the achievement of the National Park Service mission. The series provides contributors with a forum for displaying comprehensive data that are often deleted from journals because of page limitations. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner.