Inter-Sarcomere Coordination in Muscle Revealed Through Individual Sarcomere Response to Quick Stretch
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Microanatomy of Muscles
Microanatomy of Muscles Anatomy & Physiology Class Three Main Muscle Types Objectives: By the end of this presentation you will have the information to: 1. Describe the 3 main types of muscles. 2. Detail the functions of the muscle system. 3. Correctly label the parts of a myocyte (muscle cell) 4. Identify the levels of organization in a skeletal muscle from organ to myosin. 5. Explain how a muscle contracts utilizing the correct terminology of the sliding filament theory. 6. Contrast and compare cardiac and smooth muscle with skeletal muscle. Major Functions: Muscle System 1. Moving the skeletal system and posture. 2. Passing food through the digestive system & constriction of other internal organs. 3. Production of body heat. 4. Pumping the blood throughout the body. 5. Communication - writing and verbal Specialized Cells (Myocytes) ~ Contractile Cells Can shorten along one or more planes because of specialized cell membrane (sarcolemma) and specialized cytoskeleton. Specialized Structures found in Myocytes Sarcolemma: The cell membrane of a muscle cell Transverse tubule: a tubular invagination of the sarcolemma of skeletal or cardiac muscle fibers that surrounds myofibrils; involved in transmitting the action potential from the sarcolemma to the interior of the myofibril. Sarcoplasmic Reticulum: The special type of smooth endoplasmic Myofibrils: reticulum found in smooth and a contractile fibril of skeletal muscle, composed striated muscle fibers whose function mainly of actin and myosin is to store and release calcium ions. Multiple Nuclei (skeletal) & many mitochondria Skeletal Muscle - Microscopic Anatomy A whole skeletal muscle (such as the biceps brachii) is considered an organ of the muscular system. Each organ consists of skeletal muscle tissue, connective tissue, nerve tissue, and blood or vascular tissue. -

Vocabulario De Morfoloxía, Anatomía E Citoloxía Veterinaria
Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) Servizo de Normalización Lingüística Universidade de Santiago de Compostela COLECCIÓN VOCABULARIOS TEMÁTICOS N.º 4 SERVIZO DE NORMALIZACIÓN LINGÜÍSTICA Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) 2008 UNIVERSIDADE DE SANTIAGO DE COMPOSTELA VOCABULARIO de morfoloxía, anatomía e citoloxía veterinaria : (galego-español- inglés) / coordinador Xusto A. Rodríguez Río, Servizo de Normalización Lingüística ; autores Matilde Lombardero Fernández ... [et al.]. – Santiago de Compostela : Universidade de Santiago de Compostela, Servizo de Publicacións e Intercambio Científico, 2008. – 369 p. ; 21 cm. – (Vocabularios temáticos ; 4). - D.L. C 2458-2008. – ISBN 978-84-9887-018-3 1.Medicina �������������������������������������������������������������������������veterinaria-Diccionarios�������������������������������������������������. 2.Galego (Lingua)-Glosarios, vocabularios, etc. políglotas. I.Lombardero Fernández, Matilde. II.Rodríguez Rio, Xusto A. coord. III. Universidade de Santiago de Compostela. Servizo de Normalización Lingüística, coord. IV.Universidade de Santiago de Compostela. Servizo de Publicacións e Intercambio Científico, ed. V.Serie. 591.4(038)=699=60=20 Coordinador Xusto A. Rodríguez Río (Área de Terminoloxía. Servizo de Normalización Lingüística. Universidade de Santiago de Compostela) Autoras/res Matilde Lombardero Fernández (doutora en Veterinaria e profesora do Departamento de Anatomía e Produción Animal. -

(7E) Powerpoint Lecture Outline Chapter 8: Control of Movement
Carlson (7e) PowerPoint Lecture Outline Chapter 8: Control of Movement This multimedia product and its contents are protected under copyright law. The following are prohibited by law: •any public performance or display, including transmission of any image over a network; •preparation of any derivative work, including extraction, in whole or in part, of any images; •any rental, lease, or lending of the program. Copyright 2001 by Allyn & Bacon Skeletal Muscle n Movements of our body are accomplished by contraction of the skeletal muscles l Flexion: contraction of a flexor muscle draws in a limb l Extension: contraction of extensor muscle n Skeletal muscle fibers have a striated appearance n Skeletal muscle is composed of two fiber types: l Extrafusal: innervated by alpha-motoneurons from the spinal cord: exert force l Intrafusal: sensory fibers that detect stretch of the muscle u Afferent fibers: report length of intrafusal: when stretched, the fibers stimulate the alpha-neuron that innervates the muscle fiber: maintains muscle tone u Efferent fibers: contraction adjusts sensitivity of afferent fibers. 8.2 Copyright 2001 by Allyn & Bacon Skeletal Muscle Anatomy n Each muscle fiber consists of a bundle of myofibrils l Each myofibril is made up of overlapping strands of actin and myosin l During a muscle twitch, the myosin filaments move relative to the actin filaments, thereby shortening the muscle fiber 8.3 Copyright 2001 by Allyn & Bacon Neuromuscular Junction n The neuromuscular junction is the synapse formed between an alpha motor neuron -

CAP2 Deficiency Delays Myofibril Actin Cytoskeleton Differentiation and Disturbs Skeletal Muscle Architecture and Function
CAP2 deficiency delays myofibril actin cytoskeleton differentiation and disturbs skeletal muscle architecture and function Lara-Jane Kepsera, Fidan Damara, Teresa De Ciccob, Christine Chaponnierc, Tomasz J. Prószynski b, Axel Pagenstecherd, and Marco B. Rusta,e,f,1 aMolecular Neurobiology Group, Institute of Physiological Chemistry, University of Marburg, 35032 Marburg, Germany; bLaboratory of Synaptogenesis, Nencki Institute of Experimental Biology PAS, 02-093 Warsaw, Poland; cDepartment of Pathology and Immunology, University of Geneva, 1211 Geneva, Switzerland; dInstitute of Neuropathology, University of Marburg, 35032 Marburg, Germany; eCenter for Mind, Brain and Behavior, Research Campus of Central Hessen, 35032 Marburg, Germany; and fDFG Research Training Group “Membrane Plasticity in Tissue Development and Remodeling,” GRK 2213, University of Marburg, 35032 Marburg, Germany Edited by Yale E. Goldman, University of Pennsylvania/PMI, Philadelphia, PA, and approved March 14, 2019 (received for review August 7, 2018) Actin filaments (F-actin) are key components of sarcomeres, the have acquired specific functions. While previous analyses of mu- basic contractile units of skeletal muscle myofibrils. A crucial step tant mice demonstrated a role of CAP2 in neuron morphology and during myofibril differentiation is the sequential exchange of heart physiology (13–15), its function in skeletal muscles has not α-actin isoforms from smooth muscle (α-SMA) and cardiac (α-CAA) been investigated, yet. to skeletal muscle α-actin (α-SKA) that, in mice, occurs during early We here report a function for CAP2 in skeletal muscle de- postnatal life. This “α-actin switch” requires the coordinated activ- velopment. We found that CAP2 controls the exchange of ity of actin regulators because it is vital that sarcomere structure α-actin isoforms during myofibril differentiation. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

Breakdown of Filamentous Myofibrils by the UPS–Step by Step
biomolecules Review Breakdown of Filamentous Myofibrils by the UPS–Step by Step Dina Aweida and Shenhav Cohen * Faculty of Biology, Technion Institute of Technology, Haifa 32000, Israel; [email protected] * Correspondence: [email protected]; Tel.: +972-4-8294214 Abstract: Protein degradation maintains cellular integrity by regulating virtually all biological pro- cesses, whereas impaired proteolysis perturbs protein quality control, and often leads to human disease. Two major proteolytic systems are responsible for protein breakdown in all cells: autophagy, which facilitates the loss of organelles, protein aggregates, and cell surface proteins; and the ubiquitin- proteasome system (UPS), which promotes degradation of mainly soluble proteins. Recent findings indicate that more complex protein structures, such as filamentous assemblies, which are not acces- sible to the catalytic core of the proteasome in vitro, can be efficiently degraded by this proteolytic machinery in systemic catabolic states in vivo. Mechanisms that loosen the filamentous structure seem to be activated first, hence increasing the accessibility of protein constituents to the UPS. In this review, we will discuss the mechanisms underlying the disassembly and loss of the intricate insoluble filamentous myofibrils, which are responsible for muscle contraction, and whose degradation by the UPS causes weakness and disability in aging and disease. Several lines of evidence indicate that myofibril breakdown occurs in a strictly ordered and controlled manner, and the function of AAA-ATPases is crucial for their disassembly and loss. Keywords: ubiquitin; proteasome; autophagy; muscle atrophy; intermediate filaments; myofibrils; AAA-ATPases 1. Introduction Citation: Aweida, D.; Cohen, S. Proteolysis promotes tissue homeostasis by controlling protein abundance in response Breakdown of Filamentous Myofibrils to extracellular and intracellular cues, and by preventing accumulation of misfolded or by the UPS–Step by Step. -

Microfluidic Perfusion Shows Intersarcomere Dynamics Within Single Skeletal Muscle Myofibrils
Microfluidic perfusion shows intersarcomere dynamics within single skeletal muscle myofibrils Felipe de Souza Leitea, Fabio C. Minozzoa, David Altmanb, and Dilson E. Rassiera,c,d,1 aDepartment of Kinesiology and Physical Education, McGill University, Montreal, QC, Canada, H2W 1S4; bDepartment of Physics, Willamette University, Salem, OR 97301; cDepartment of Physiology, McGill University, Montreal, QC, Canada, H3G 1Y6; and dDepartment of Physics, McGill University, Montreal, QC, Canada H3A 2T8 Edited by James A. Spudich, Stanford University School of Medicine, Stanford, CA, and approved July 5, 2017 (received for review January 30, 2017) The sarcomere is the smallest functional unit of myofibrils in passive force production, which enables the myofibrils to stabilize in striated muscles. Sarcomeres are connected in series through a a given activation condition. Because of technical limitations, the network of elastic and structural proteins. During myofibril activa- mechanisms governing the interactionofsarcomeresinamyofibril tion, sarcomeres develop forces that are regulated through complex and its consequences for force production remain unclear (11). dynamics among their structures. The mechanisms that regulate In this study, we tested the hypotheses that the mechanical work intersarcomere dynamics are unclear, which limits our understand- of one sarcomere effectively communicates with other sarcomeres ing of fundamental muscle features. Such dynamics are associated in series through the passive work of titin and that myofibril me- with the loss in forces caused by mechanical instability encountered chanics are largely governed by intersarcomere dynamics. To test in muscle diseases and cardiomyopathy and may underlie potential these hypotheses, we used microfluidic perfusions to locally control target treatments for such conditions. In this study, we developed a one sarcomere or a predetermined group of sarcomeres within an microfluidic perfusion system to control one sarcomere within a isolated myofibril. -

Muscle Contraction
10/19/2009 CONTROL OF MOVEMENT: STRIATED MUSCLES SKELETAL (STRIATED) MUSCLE: - each muscle = ____________________________________ - each muscle cell = _______________________________________ -Myosin: Filamentous___________________________________________ protein with cross bridges -Actin: _________________________________________________Filamentous protein where cross bridges of myosin bind ANATOMY OF SKELETAL MUSCLE ______________ _____________ ________ Extrafusal muscle fiber ________ _________ Myofibril _____________ __________________ ___________ MUSCLE CONTRACTION Watch muscle contraction movie Myosin Myosin cross bridges filament Actin filaments Actin MtfMovement of filament actin filament Myosin cross bridge Movement of myosin filament Heads of cross bridges: 1. Attach to active sites on actin filaments 2. “Ratchet” forward 3. Release 4. Repeat -Onlyoccurs in the presence of ________ How is calcium released? From___________________________ activity at neuromuscular ________junction 1 10/19/2009 NEUROMUSCULAR JUNCTION Synapse between terminal of _________________ and a ___________ is called a neuromuscular junction; Terminals of alpha motor neurons synapse on _____________- grooves along the surface of muscle fibers; When motor neuron fires, _____________ is liberated from terminals at the endplate and depolarizes muscle fibers - ________________; Depolarization of muscle fiber opens ________ _________________________, producing a large calcium influx into the fiber; Calcium triggers the actin-myosin “rowing” action leading to the -

Proteins of the Myofibril
131. PROTEINS OF THE MYUFlBRIL A. G. SZEWT-GYORGYI Ladies and gentlemen, I would like to review here a number of questions which involve some properties of the fibrous muscle proteins, how these properties lead to contractions, what the changes are which occur during contractions, and also to expand on the paper presented by Dr. Cassens by discussing how some of these proteins are responsible for the specialized structure of muscle and what reactions may control the forma- tion of these structures and activities. The muscle proteins can be conveniently divided into different fractions according to solubility and also according to function. If you grind up muscle and add a solvent of low ionic strength you will find that 35% of the proteins will be readily solubilized without, as I think will be shown on the first slide, really disrupting the specialized structure of the muscle. The striated pattern that apgeass in unextracted muscle, as shown by Dr. Cassens, will remain. What is solubilized is the enzymes of the glycolytic cycle, the phosphate-producing enzymes like myokinase and creetinekinase, and really the residue, that is, the insoluble portion remaining, will show the structural regularity of the muscle even clearer than before the extraction, We do not know exactly where these soluble enzymes are localized or if they are localized, but we think they are somewhere in the sarcoplasm and that they are not associated with the filamentous structure of muscle. I will not discuss these proteins any more. They do not concern us nor have anything to do with the culinary aspects of meat. -
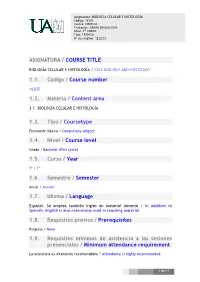
ASIGNATURA / COURSE TITLE 1.1. Código / Course Number 1.2
Asignatura: BIOLOGÍA CELULAR E HISTOLOGÍA Código: 16305 Centro: CIENCIAS Titulación: GRADO EN BIOLOGÍA Nivel: 1er CURSO Tipo: TRONCAL Nº de créditos: 12 ECTS ASIGNATURA / COURSE TITLE BIOLOGÍA CELULAR E HISTOLOGÍA / CELL BIOLOGY AND HISTOLOGY 1.1. Código / Course number 16305 1.2. Materia / Content area 3.1. BIOLOGÍA CELULAR E HISTOLOGÍA 1.3. Tipo / Coursetype Formación básica / Compulsory subject 1.4. Nivel / Course level Grado / Bachelor (first cycle) 1.5. Curso / Year 1º / 1st 1.6. Semestre / Semester Anual / Annual 1.7. Idioma / Language Español. Se emplea también Inglés en material docente / In addition to Spanish, English is also extensively used in teaching material 1.8. Requisitos previos / Prerequisites Ninguno / None 1.9. Requisitos mínimos de asistencia a las sesiones presenciales / Minimum attendance requirement La asistencia es altamente recomendable / Attendance is highly recommended 1 de 17 Asignatura: BIOLOGÍA CELULAR E HISTOLOGÍA Código: 16305 Centro: CIENCIAS Titulación: GRADO EN BIOLOGÍA Nivel: 1er CURSO Tipo: TRONCAL Nº de créditos: 12 ECTS 1.10. Datos del equipo docente /Faculty data Los profesores implicados en esta asignatura serán todos los del área de Biología Celular del Departamento de Biología de la U.A.M. El coordinador de la asignatura será: All the teachers of the Cell Biology Area, from the Biology Department of UAM, will be involved in this course. The coordinator of the course will be: Coordinador: Profesor/a: ALBERTO VIERA VICARIO Correo electrónico: Teléfono: Web del profesor: [email protected] 914978914 Departamento: BIOLOGÍA Centro: FACULTAD DE CIENCIAS Horario de tutorías generales: DE LUNES A VIERNES PREVIA CITA Despacho: A-107 1.11. -

The Unified Myofibrillar Matrix for Force Generation in Muscle
ARTICLE https://doi.org/10.1038/s41467-020-17579-6 OPEN The unified myofibrillar matrix for force generation in muscle ✉ T. Bradley Willingham1, Yuho Kim 1, Eric Lindberg1, Christopher K. E. Bleck 1 & Brian Glancy 1,2 Human movement occurs through contraction of the basic unit of the muscle cell, the sarcomere. Sarcomeres have long been considered to be arranged end-to-end in series along the length of the muscle into tube-like myofibrils with many individual, parallel myofibrils 1234567890():,; comprising the bulk of the muscle cell volume. Here, we demonstrate that striated muscle cells form a continuous myofibrillar matrix linked together by frequently branching sarco- meres. We find that all muscle cells contain highly connected myofibrillar networks though the frequency of sarcomere branching goes down from early to late postnatal development and is higher in slow-twitch than fast-twitch mature muscles. Moreover, we show that the myofibrillar matrix is united across the entire width of the muscle cell both at birth and in mature muscle. We propose that striated muscle force is generated by a singular, mesh-like myofibrillar network rather than many individual, parallel myofibrils. 1 National Heart, Lung and Blood Institute, National Institutes of Health, Bethesda, MD 20892, USA. 2 National Institute of Arthritis and Musculoskeletal and ✉ Skin Diseases, National Institutes of Health, Bethesda, MD 20892, USA. email: [email protected] NATURE COMMUNICATIONS | (2020) 11:3722 | https://doi.org/10.1038/s41467-020-17579-6 | www.nature.com/naturecommunications 1 ARTICLE NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-020-17579-6 he mechanisms underlying how muscle contraction gen- muscle cell structures for abnormalities. -

The Role of Titin and Nebulin in Myofibril Assembly in Cultured Embryonic Chick Muscle Cells
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1988 The oler of titin and nebulin in myofibril assembly in cultured embryonic chick muscle cells Michelle Ann Kurpakus Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Biochemistry Commons Recommended Citation Kurpakus, Michelle Ann, "The or le of titin and nebulin in myofibril assembly in cultured embryonic chick muscle cells " (1988). Retrospective Theses and Dissertations. 9689. https://lib.dr.iastate.edu/rtd/9689 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS The most advanced technology has been used to photo graph and reproduce this manuscript from the microfilm master. UMI films the original text directly firom the copy submitted. Thus, some dissertation copies are in typewriter face, while others may be from a computer printer. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyrighted material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are re produced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. Each oversize page is available as one exposure on a standard 35 mm slide or as a 17" x 23" black and white photographic print for an additional charge.