Vitales, C Nymphaeales Austrobaileyales
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Insights Into Tyrosinase Inhibition by Compounds Isolated from Greyia Radlkoferi Syzsyl Using Biological Activity, Molecular Docking and Gene Expression Analysis
Insights into tyrosinase inhibition by compounds isolated from Greyia radlkoferi Syzsyl using biological activity, molecular docking and gene expression analysis Namrita Lall a *, Elizabeth Mogapi a, Marco Nuno de Canha a, Bridget Crampton a, Mabatho Nqephe a, Ahmed Hussein a, b and Vivek Kumara a Department of Plant and Soil Sciences, University of Pretoria, Pretoria, South Africa, 0002 b Department of Chemistry, University of Western Cape, Private Bag X17, Bellville, 7535, Cape Town Corresponding author: Professor Namrita Lall Department of Plant and Soil Sciences Plant Science Complex, University of Pretoria Pretoria, South Africa, 0002 Email: [email protected] Tel.: 00 27 12 420 2524 Fax: 00 27 12 420 6668 1 ABSTRACT Greyia radlkoferi ethanol extract and its five compounds were tested for their inhibitory activity against the mushroom tyrosinase enzyme and melanin production on melanocytes. The crude extract showed significant tyrosinase inhibition with IC50 of 17.96µg/ml. This is the first report of the isolation of these 5 compounds from Greyia radlkoferi. 2’, 4’, 6’- trihydroxydihydrochalcone showed the highest tyrosinase inhibition at 17.70µg/ml (68.48µM), with low toxicity when compared with crude extract. This compound is therefore, a key component in the crude extract, which is responsible for tyrosinase inhibitory activity. The RT-qPCR indicated that the mechanism of action is most likely post transcriptional. Further, the molecular docking study showed that tyrosinase inhibitory activity depends on interaction of the compound with Cu2+ ions at the active site. This is the first report of the tyrosinase inhibitory activity of the G. radlkoferi extract and molecular insights on interaction of its compounds with Cu2+ ions as the driving factor for tyrosinase inhibition. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Calibrated Chronograms, Fossils, Outgroup Relationships, and Root Priors: Re-Examining the Historical Biogeography of Geraniales
bs_bs_banner Biological Journal of the Linnean Society, 2014, 113, 29–49. With 4 figures Calibrated chronograms, fossils, outgroup relationships, and root priors: re-examining the historical biogeography of Geraniales KENNETH J. SYTSMA1,*, DANIEL SPALINK1 and BRENT BERGER2 1Department of Botany, University of Wisconsin, Madison, WI 53706, USA 2Department of Biological Sciences, St. John’s University, Queens, NY 11439, USA Received 26 November 2013; revised 23 February 2014; accepted for publication 24 February 2014 We re-examined the recent study by Palazzesi et al., (2012) published in the Biological Journal of the Linnean Society (107: 67–85), that presented the historical diversification of Geraniales using BEAST analysis of the plastid spacer trnL–F and of the non-coding nuclear ribosomal internal transcribed spacers (ITS). Their study presented a set of new fossils within the order, generated a chronogram for Geraniales and other rosid orders using fossil-based priors on five nodes, demonstrated an Eocene radiation of Geraniales (and other rosid orders), and argued for more recent (Pliocene–Pleistocene) and climate-linked diversification of genera in the five recognized families relative to previous studies. As a result of very young ages for the crown of Geraniales and other rosid orders, unusual relationships of Geraniales to other rosids, and apparent nucleotide substitution saturation of the two gene regions, we conducted a broad series of BEAST analyses that incorporated additional rosid fossil priors, used more accepted rosid ordinal -

Literature Cited
Literature Cited Robert W. Kiger, Editor This is a consolidated list of all works cited in volume 9, whether as selected references, in text, or in nomenclatural contexts. In citations of articles, both here and in the taxonomic treatments, and also in nomenclatural citations, the titles of serials are rendered in the forms recommended in G. D. R. Bridson and E. R. Smith (1991), Bridson (2004), and Bridson and D. W. Brown (http://fmhibd.library.cmu.edu/fmi/iwp/cgi?-db=BPH_Online&-loadframes). When those forms are abbreviated, as most are, cross references to the corresponding full serial titles are interpolated here alphabetically by abbreviated form. In nomenclatural citations (only), book titles are rendered in the abbreviated forms recommended in F. A. Stafleu and R. S. Cowan (1976–1988) and Stafleu et al. (1992–2009). Here, those abbreviated forms are indicated parenthetically following the full citations of the corresponding works, and cross references to the full citations are interpolated in the list alphabetically by abbreviated form. Two or more works published in the same year by the same author or group of coauthors will be distinguished uniquely and consistently throughout all volumes of Flora of North America by lower-case letters (b, c, d, ...) suffixed to the date for the second and subsequent works in the set. The suffixes are assigned in order of editorial encounter and do not reflect chronological sequence of publication. The first work by any particular author or group from any given year carries the implicit date suffix “a”; thus, the sequence of explicit suffixes begins with “b”. -

Phylogeny and Historical Biogeography of Geraniaceae In
Systematic Botany (2008), 33(2): pp. 326–342 © Copyright 2008 by the American Society of Plant Taxonomists Phylogeny and Historical Biogeography of Geraniaceae in Relation to Climate Changes and Pollination Ecology Omar Fiz, Pablo Vargas, Marisa Alarcón, Carlos Aedo, José Luis García, and Juan José Aldasoro1 Real Jardín Botanico de Madrid, CSIC, Plaza de Murillo 2, 28014 Madrid, Spain 1Author for correspondence ([email protected]) Communicating Editor: Mark P. Simmons Abstract—Chloroplast (trnL–F and rbcL) sequences were used to reconstruct the phylogeny of Geraniaceae and Hypseocharitaceae. According to these data Hypseocharitaceae and Geraniaceae are monophyletic. Pelargonium and Monsonia are sisters to the largest clade of Geraniaceae, formed by Geranium, Erodium and California. According to molecular dating and dispersal-vicariance analysis, the split of the stem branches of Geraniaceae probably occurred during the Oligocene, in southern Africa or in southern Africa plus the Mediterranean area. However, their diversification occurred during the Miocene, coinciding with the beginning of major aridification events in their distribution areas. An ancestor of the largest clade of Geraniaceae (Geranium, Erodium, and California) colonised a number of habitats in the northern hemisphere and in South American mountain ranges. In summary, the evolution of the Geraniaceae is marked by the dispersal of ancestors from Southern Africa to cold, temperate and often disturbed habitats in the rest of world, where only generalist pollination and facultative autogamy could ensure sufficient seed production and survival. Keywords—autocompatibility, dispersal-vicariance, drought-tolerance, molecular dating, nectaries, P/O indexes. The Geraniaceae are included in the order Geraniales along are characteristic of the Afro-Arabian land mass (Hutchin- with the families Francoaceae, Greyiaceae, Ledocarpaceae, son 1969). -

Vascular Plants of Eagle Creek Campground, Trinity County
Humboldt State University Digital Commons @ Humboldt State University Botanical Studies Open Educational Resources and Data 9-17-2018 Vascular Plants of Eagle Creek Campground, Trinity County James P. Smith Jr Humboldt State University, [email protected] Follow this and additional works at: https://digitalcommons.humboldt.edu/botany_jps Part of the Botany Commons Recommended Citation Smith, James P. Jr, "Vascular Plants of Eagle Creek Campground, Trinity County" (2018). Botanical Studies. 84. https://digitalcommons.humboldt.edu/botany_jps/84 This Flora of Northwest California-Checklists of Local Sites is brought to you for free and open access by the Open Educational Resources and Data at Digital Commons @ Humboldt State University. It has been accepted for inclusion in Botanical Studies by an authorized administrator of Digital Commons @ Humboldt State University. For more information, please contact [email protected]. THE VASCULAR PLANTS OF EAGLE CREEK CAMPGROUND (TRINITY COUNTY, CALIFORNIA) Compiled by James P. Smith, Jr. Professor Emeritus of Botany Department of Biological Sciences Humboldt State University Fifth Edition • 14 September 2018 The Eagle Creek Campground is located about Boraginaceae five miles north of Coffee Creek along State Route Amsinckia intermedia 3 in the Shasta-Trinity National Forest. On one Lithospermum californicum side is the Trinity River and Eagle Creek on the Plagiobothrys tenellus other. Caryophyllaceae F E R N S Eremogone congesta Silene lemmonii Equisetum hyemale var. affine Stellaria media Cheilanthes siliquosa Stellaria nitens Polystichum imbricans ssp. imbricans Pteridium aquilinum var. pubescens Compositae (Asteraceae) Adenocaulon bicolor C O N I F E R S Ageratina occidentale Agoseris heterophylla Abies concolor Antennaria argentea Calocedrus decurrens Antennaria dimorpha Pinus jeffreyi Balsamorhiza deltoidea Pinus ponderosa var. -

Washington Flora Checklist a Checklist of the Vascular Plants of Washington State Hosted by the University of Washington Herbarium
Washington Flora Checklist A checklist of the Vascular Plants of Washington State Hosted by the University of Washington Herbarium The Washington Flora Checklist aims to be a complete list of the native and naturalized vascular plants of Washington State, with current classifications, nomenclature and synonymy. The checklist currently contains 3,929 terminal taxa (species, subspecies, and varieties). Taxa included in the checklist: * Native taxa whether extant, extirpated, or extinct. * Exotic taxa that are naturalized, escaped from cultivation, or persisting wild. * Waifs (e.g., ballast plants, escaped crop plants) and other scarcely collected exotics. * Interspecific hybrids that are frequent or self-maintaining. * Some unnamed taxa in the process of being described. Family classifications follow APG IV for angiosperms, PPG I (J. Syst. Evol. 54:563?603. 2016.) for pteridophytes, and Christenhusz et al. (Phytotaxa 19:55?70. 2011.) for gymnosperms, with a few exceptions. Nomenclature and synonymy at the rank of genus and below follows the 2nd Edition of the Flora of the Pacific Northwest except where superceded by new information. Accepted names are indicated with blue font; synonyms with black font. Native species and infraspecies are marked with boldface font. Please note: This is a working checklist, continuously updated. Use it at your discretion. Created from the Washington Flora Checklist Database on September 17th, 2018 at 9:47pm PST. Available online at http://biology.burke.washington.edu/waflora/checklist.php Comments and questions should be addressed to the checklist administrators: David Giblin ([email protected]) Peter Zika ([email protected]) Suggested citation: Weinmann, F., P.F. Zika, D.E. Giblin, B. -

Parallel Evolution of Highly Conserved Plastid Genome Architecture in Red Seaweeds and Seed Plants
Lee et al. BMC Biology (2016) 14:75 DOI 10.1186/s12915-016-0299-5 RESEARCH ARTICLE Open Access Parallel evolution of highly conserved plastid genome architecture in red seaweeds and seed plants JunMo Lee1, Chung Hyun Cho1, Seung In Park1, Ji Won Choi1, Hyun Suk Song1, John A. West2, Debashish Bhattacharya3† and Hwan Su Yoon1*† Abstract Background: The red algae (Rhodophyta) diverged from the green algae and plants (Viridiplantae) over one billion years ago within the kingdom Archaeplastida. These photosynthetic lineages provide an ideal model to study plastid genome reduction in deep time. To this end, we assembled a large dataset of the plastid genomes that were available, including 48 from the red algae (17 complete and three partial genomes produced for this analysis) to elucidate the evolutionary history of these organelles. Results: We found extreme conservation of plastid genome architecture in the major lineages of the multicellular Florideophyceae red algae. Only three minor structural types were detected in this group, which are explained by recombination events of the duplicated rDNA operons. A similar high level of structural conservation (although with different gene content) was found in seed plants. Three major plastid genome architectures were identified in representatives of 46 orders of angiosperms and three orders of gymnosperms. Conclusions: Our results provide a comprehensive account of plastid gene loss and rearrangement events involving genome architecture within Archaeplastida and lead to one over-arching conclusion: from an ancestral pool of highly rearranged plastid genomes in red and green algae, the aquatic (Florideophyceae) and terrestrial (seed plants) multicellular lineages display high conservation in plastid genome architecture. -

Staminodes: Their Morphological and Evolutionary Significance Author(S): L
Staminodes: Their Morphological and Evolutionary Significance Author(s): L. P. Ronse Decraene and E. F. Smets Source: Botanical Review, Vol. 67, No. 3 (Jul. - Sep., 2001), pp. 351-402 Published by: Springer on behalf of New York Botanical Garden Press Stable URL: http://www.jstor.org/stable/4354395 . Accessed: 23/06/2014 03:18 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. New York Botanical Garden Press and Springer are collaborating with JSTOR to digitize, preserve and extend access to Botanical Review. http://www.jstor.org This content downloaded from 210.72.93.185 on Mon, 23 Jun 2014 03:18:32 AM All use subject to JSTOR Terms and Conditions THE BOTANICAL REVIEW VOL. 67 JULY-SEPTEMBER 2001 No. 3 Staminodes: Their Morphological and Evolutionary Signiflcance L. P. RONSEDECRAENE AND E. F. SMETS Katholieke UniversiteitLeuven Laboratory of Plant Systematics Institutefor Botany and Microbiology KasteelparkArenberg 31 B-3001 Leuven, Belgium I. Abstract........................................... 351 II. Introduction.................................................... 352 III. PossibleOrigin of Staminodes........................................... 354 IV. A Redefinitionof StaminodialStructures .................................. 359 A. Surveyof the Problem:Case Studies .............. .................... 359 B. Evolutionof StaminodialStructures: Function-Based Definition ... ......... 367 1. VestigialStaminodes ........................................... 367 2. FunctionalStaminodes ........................................... 368 C. StructuralSignificance of StaminodialStructures: Topology-Based Definition . -

Annotated Checklist of the Vascular Plant Flora of Grand Canyon-Parashant National Monument Phase II Report
Annotated Checklist of the Vascular Plant Flora of Grand Canyon-Parashant National Monument Phase II Report By Dr. Terri Hildebrand Southern Utah University, Cedar City, UT and Dr. Walter Fertig Moenave Botanical Consulting, Kanab, UT Colorado Plateau Cooperative Ecosystems Studies Unit Agreement # H1200-09-0005 1 May 2012 Prepared for Grand Canyon-Parashant National Monument Southern Utah University National Park Service Mojave Network TABLE OF CONTENTS Page # Introduction . 4 Study Area . 6 History and Setting . 6 Geology and Associated Ecoregions . 6 Soils and Climate . 7 Vegetation . 10 Previous Botanical Studies . 11 Methods . 17 Results . 21 Discussion . 28 Conclusions . 32 Acknowledgments . 33 Literature Cited . 34 Figures Figure 1. Location of Grand Canyon-Parashant National Monument in northern Arizona . 5 Figure 2. Ecoregions and 2010-2011 collection sites in Grand Canyon-Parashant National Monument in northern Arizona . 8 Figure 3. Soil types and 2010-2011 collection sites in Grand Canyon-Parashant National Monument in northern Arizona . 9 Figure 4. Increase in the number of plant taxa confirmed as present in Grand Canyon- Parashant National Monument by decade, 1900-2011 . 13 Figure 5. Southern Utah University students enrolled in the 2010 Plant Anatomy and Diversity course that collected during the 30 August 2010 experiential learning event . 18 Figure 6. 2010-2011 collection sites and transportation routes in Grand Canyon-Parashant National Monument in northern Arizona . 22 2 TABLE OF CONTENTS Page # Tables Table 1. Chronology of plant-collecting efforts at Grand Canyon-Parashant National Monument . 14 Table 2. Data fields in the annotated checklist of the flora of Grand Canyon-Parashant National Monument (Appendices A, B, C, and D) . -

Vascular Plant Species of the Comanche National Grassland in United States Department Southeastern Colorado of Agriculture
Vascular Plant Species of the Comanche National Grassland in United States Department Southeastern Colorado of Agriculture Forest Service Donald L. Hazlett Rocky Mountain Research Station General Technical Report RMRS-GTR-130 June 2004 Hazlett, Donald L. 2004. Vascular plant species of the Comanche National Grassland in southeast- ern Colorado. Gen. Tech. Rep. RMRS-GTR-130. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 36 p. Abstract This checklist has 785 species and 801 taxa (for taxa, the varieties and subspecies are included in the count) in 90 plant families. The most common plant families are the grasses (Poaceae) and the sunflower family (Asteraceae). Of this total, 513 taxa are definitely known to occur on the Comanche National Grassland. The remaining 288 taxa occur in nearby areas of southeastern Colorado and may be discovered on the Comanche National Grassland. The Author Dr. Donald L. Hazlett has worked as an ecologist, botanist, ethnobotanist, and teacher in Latin America and in Colorado. He has specialized in the flora of the eastern plains since 1985. His many years in Latin America prompted him to include Spanish common names in this report, names that are seldom reported in floristic pub- lications. He is also compiling plant folklore stories for Great Plains plants. Since Don is a native of Otero county, this project was of special interest. All Photos by the Author Cover: Purgatoire Canyon, Comanche National Grassland You may order additional copies of this publication by sending your mailing information in label form through one of the following media. -
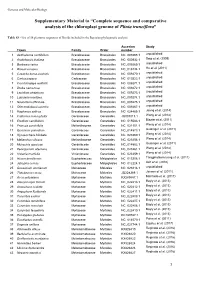
Complete Sequence and Comparative Analysis of the Chloroplast Genome of Plinia Trunciflora”
Genetics and Molecular Biology Supplementary Material to “Complete sequence and comparative analysis of the chloroplast genome of Plinia trunciflora” Table S3 - List of 56 plastome sequences of Rosids included in the Bayesian phylogenetic analysis. Accesion Study Taxon Family Order number 1 Aethionema cordifolium Brassicaceae Brassicales NC_009265.1 unpublished 2 Arabidopsis thaliana Brassicaceae Brassicales NC_000932.1 Sato et al. (1999) 3 Barbarea verna Brassicaceae Brassicales NC_009269.1 unpublished 4 Brassica napus Brassicaceae Brassicales NC_016734.1 Hu et al. (2011) 5 Capsella bursa-pastoris Brassicaceae Brassicales NC_009270.1 unpublished 6 Carica papaya Caricaceae Brassicales NC_010323.1 unpublished 7 Crucihimalaya wallichii Brassicaceae Brassicales NC_009271.1 unpublished 8 Draba nemorosa Brassicaceae Brassicales NC_009272.1 unpublished 9 Lepidium virginicum Brassicaceae Brassicales NC_009273.1 unpublished 10 Lobularia maritima Brassicaceae Brassicales NC_009274.1 unpublished 11 Nasturtium officinale Brassicaceae Brassicales NC_009275.1 unpublished 12 Olimarabidopsis pumila Brassicaceae Brassicales NC_009267.1 unpublished 13 Raphanus sativus Brassicaceae Brassicales NC_024469.1 Jeong et al. (2014) 14 California macrophylla Geraniaceae Geraniales JQ031013.1 Weng et al. (2014) 15 Erodium carvifolium Geraniaceae Geraniales NC_015083.1 Blazier et al. (2011) 16 Francoa sonchifolia Melianthaceae Geraniales NC_021101.1 Weng et al. (2014) 17 Geranium palmatum Geraniaceae Geraniales NC_014573.1 Guisinger et al. (2011) 18 Hypseocharis bilobate Geraniaceae Geraniales NC_023260.1 Weng et al. (2014) 19 Melianthus villosus Melianthaceae Geraniales NC_023256.1 Weng et al. (2014) 20 Monsonia speciose Geraniaceae Geraniales NC_014582.1 Guisinger et al. (2011) 21 Pelargonium alternans Geraniaceae Geraniales NC_023261.1 Weng et al. (2014) 22 Viviania marifolia Vivianiaceae Geraniales NC_023259.1 Weng et al. (2014) 23 Hevea brasiliensis Euphorbiaceae Malpighiales NC_015308.1 Tangphatsornruang et al.