Drivers of Airborne Human-To-Human Pathogen Transmission
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Human Illness Caused by E. Coli O157:H7 from Food and Non-Food Sources
FRI BRIEFINGS Human Illness Caused by E. coli O157:H7 from Food and Non-food Sources M. Ellin Doyle1*, John Archer2, Charles W. Kaspar1, and Ronald Weiss1 1Food Research Institute, University of Wisconsin–Madison, Madison, WI 53706 2Wisconsin Division of Public Health, Bureau of Communicable Diseases and Preparedness, Communicable Disease Epidemiology Section, Madison, WI 53702 Contents Introduction ...................................................................................................................................2 Epidemiology of E. coli O157:H7..................................................................................................2 Outbreak Data ........................................................................................................................2 Reservoirs of E. coli O157:H7 ..............................................................................................3 Cattle—the primary reservoir ........................................................................................3 Other ruminants .............................................................................................................4 Other animals .................................................................................................................4 Transport Hosts......................................................................................................................4 Routes of Human Infection ....................................................................................................5 -

How Do Pathogenic Microorganisms Develop Cross-Kingdom Host Jumps? Peter Van Baarlen1, Alex Van Belkum2, Richard C
Molecular mechanisms of pathogenicity: how do pathogenic microorganisms develop cross-kingdom host jumps? Peter van Baarlen1, Alex van Belkum2, Richard C. Summerbell3, Pedro W. Crous3 & Bart P.H.J. Thomma1 1Laboratory of Phytopathology, Wageningen University, Wageningen, The Netherlands; 2Department of Medical Microbiology and Infectious Diseases, Erasmus MC, University Medical Centre Rotterdam, Rotterdam, The Netherlands; and 3CBS Fungal Biodiversity Centre, Utrecht, The Netherlands Correspondence: Bart P.H.J. Thomma, Abstract Downloaded from https://academic.oup.com/femsre/article/31/3/239/2367343 by guest on 27 September 2021 Laboratory of Phytopathology, Wageningen University, Binnenhaven 5, 6709 PD It is common knowledge that pathogenic viruses can change hosts, with avian Wageningen, The Netherlands. Tel.: 10031 influenza, the HIV, and the causal agent of variant Creutzfeldt–Jacob encephalitis 317 484536; fax: 10031 317 483412; as well-known examples. Less well known, however, is that host jumps also occur e-mail: [email protected] with more complex pathogenic microorganisms such as bacteria and fungi. In extreme cases, these host jumps even cross kingdom of life barriers. A number of Received 3 July 2006; revised 22 December requirements need to be met to enable a microorganism to cross such kingdom 2006; accepted 23 December 2006. barriers. Potential cross-kingdom pathogenic microorganisms must be able to First published online 26 February 2007. come into close and frequent contact with potential hosts, and must be able to overcome or evade host defences. Reproduction on, in, or near the new host will DOI:10.1111/j.1574-6976.2007.00065.x ensure the transmission or release of successful genotypes. -

REALM Research Briefing: Vaccines, Variants, and Venitlation
Briefing: Vaccines, Variants, and Ventilation A Briefing on Recent Scientific Literature Focused on SARS-CoV-2 Vaccines and Variants, Plus the Effects of Ventilation on Virus Spread Dates of Search: 01 January 2021 through 05 July 2021 Published: 22 July 2021 This document synthesizes various studies and data; however, the scientific understanding regarding COVID-19 is continuously evolving. This material is being provided for informational purposes only, and readers are encouraged to review federal, state, tribal, territorial, and local guidance. The authors, sponsors, and researchers are not liable for any damages resulting from use, misuse, or reliance upon this information, or any errors or omissions herein. INTRODUCTION Purpose of This Briefing • Access to the latest scientific research is critical as libraries, archives, and museums (LAMs) work to sustain modified operations during the continuing severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic. • As an emerging event, the SARS-CoV-2 pandemic continually presents new challenges and scientific questions. At present, SARS-CoV-2 vaccines and variants in the US are two critical areas of focus. The effects of ventilation-based interventions on the spread of SARS-CoV-2 are also an interest area for LAMs. This briefing provides key information and results from the latest scientific literature to help inform LAMs making decisions related to these topics. How to Use This Briefing: This briefing is intended to provide timely information about SARS-CoV-2 vaccines, variants, and ventilation to LAMs and their stakeholders. Due to the evolving nature of scientific research on these topics, the information provided here is not intended to be comprehensive or final. -

A Predictive Modelling Framework for COVID-19 Transmission to Inform the Management of Mass Events
medRxiv preprint doi: https://doi.org/10.1101/2021.05.13.21256857; this version posted May 16, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in perpetuity. It is made available under a CC-BY-NC-ND 4.0 International license . A Predictive Modelling Framework for COVID-19 Transmission to Inform the Management of Mass Events Claire Donnat Freddy Bunbury Department of Statistics Department of Plant Biology University of Chicago, Chicago, USA Carnegie Institution for Science, Stanford, USA [email protected] [email protected] Jack Kreindler Filippos T. Filippidis School of Public Health School of Public Health Imperial College, London, UK Imperial College, London, UK [email protected] [email protected] Austen El-Osta Tõnu Esko Matthew Harris School of Public Health Institute of Genomics School of Public Health Imperial College, London, UK University of Tartu, Tartu, Estonia Imperial College, London, UK [email protected] [email protected] [email protected] Abstract Modelling COVID-19 transmission at live events and public gatherings is essential to evaluate and control the probability of subsequent outbreaks. Model estimates can be used to inform event organizers about the possibility of super-spreading and the predicted efficacy of safety protocols, as well as to communicate to participants their personalised risk so that they may choose whether to attend. Yet, despite the fast-growing body of literature on COVID transmission dynamics, current risk models either neglect contextual information on vaccination rates or disease prevalence or do not attempt to quantitatively model transmission, thus limiting their potential to provide insightful estimates. -

From the COCCI Syndemic to the COVID-19 Pandemic
Scholars Insight Publishers Journal of Epidemiology and Global Health Research Research Article Open Access From the COCCI Syndemic to the COVID-19 Pandemic: A Cautionary Tale Interaction of Metabolic Syndrome, Obesity, Particulate Matter (PM), SARS-CoV-2 and the inflammatory response Clearfield M*, Gayer G, Wagner A, Stevenson T, Shubrook J and Gugliucci A Touro University College of Osteopathic Medicine, California, USA Corresponding Author: Dr. Clearfield M, Touro University Abstract College of Osteopathic Medicine, California, USA. A narrative review of the literature was conducted to determine E-mail id: [email protected] associations between cardiovascular (CV) risk factors associated with the COCCI syndemic (Cardiovascular disease as a result of the Received Date: March 15, 2021; interactions between obesity, climate change and inflammation) and Accepted Date: May 19, 2021; COVID-19. Published Date: May 21, 2021; The COCCI syndemic consists of two health conditions Publisher: Scholars Insight Online Publishers (dysmetabolic obesity and air pollution) that interact via biologic Citation: Clearfield M*, Gayer G, Wagner A, Stevenson T, pathways admixed with social, economic and ecologic drivers Shubrook J and Gugliucci A “From the COCCI Syndemic to augmenting adverse clinical outcomes in excess of either of these the COVID-19 Pandemic: A Cautionary Tale Interaction of health conditions individually. Metabolic Syndrome, Obesity, Particulate Matter (PM), SARS- The comorbidities noted with COVID-19 are in large part aligned CoV-2 and the inflammatory response”. J Epidemiol Glob Health with those traditional risk factors associated with CVD. In addition, Res. 2021; 1:102 when the traditional CV comorbidities are combined with the Copyright: ©2021 Clearfield M. -

Differences in Innate Immune Response Between Man and Mouse
Institute of Medical Physics and Biophysics Medical Department, Leipzig University Author Manuscript © 2014. This manuscript version is made available under the CC-BY-NC-ND 4.0 license http://creativecommons.org/licenses/by-nc-nd/4.0/ Published in final edited form as: Critical Reviews™ in Immunology, 2014; 34:433-54. Available at: http://dx.doi.org/ 10.1615/CritRevImmunol.2014011600. DIFFERENCES IN INNATE IMMUNE RESPONSE BETWEEN MAN AND MOUSE Josefin Zschaler 1,2,*, Denise Schlorke 1,2,* & Juergen Arnhold 1,2 1 Institute for Medical Physics and Biophysics, Medical Faculty, Leipzig University, Leipzig, Germany 2 Translational Centre for Regenerative Medicine (TRM), Leipzig University, Leipzig, Germany * Both authors contributed equally to this work. Abstract Mouse strains are frequently used to model human disease states, to test the efficiency of drugs and therapeutic principles. However, the direct translation of murine experimental data to human pathological events often fails due to sufficient differences in the organization of the immune system of both species. Here we give a short overview of the principle differences between mice and humans in defense strategies against pathogens and mechanisms involved in response to pathogenic microorganisms and other activators of the immune system. While in human blood mechanisms of immune resistance are highly prevailed, tolerance mechanisms dominate for the defense against pathogenic microorganisms in mouse blood. Further on, species-related differences of immune cells mainly involved in innate immune response as well as differences to maintain oxidative homeostasis are also considered. A number of disease scenarios in mice are critically reflected for their suitability to serve as a model for human pathologies. -

The Public Health Impact of Prion Diseases1
10 Feb 2005 13:10 AR AR238-PU26-08.tex AR238-PU26-08.sgm LaTeX2e(2002/01/18) P1: IBD 10.1146/annurev.publhealth.26.021304.144536 Annu. Rev. Public Health 2005. 26:191–212 doi: 10.1146/annurev.publhealth.26.021304.144536 Copyright ©c 2005 by Annual Reviews. All rights reserved First published online as a Review in Advance on December 8, 2004 THE PUBLIC HEALTH IMPACT OF PRION DISEASES1 Ermias D. Belay and Lawrence B. Schonberger Division of Viral and Rickettsial Diseases, National Center for Infectious Diseases, Centers for Disease Control and Prevention, Atlanta, Georgia 30333; email: [email protected] KeyWords transmissible spongiform encephalopathy, Creutzfeldt-Jakob disease, variant Creutzfeldt-Jakob disease, bovine spongiform encephalopathy, chronic wasting disease ■ Abstract Several prion disease–related human health risks from an exogenous source can be identified in the United States, including the iatrogenic transmission of Creutzfeldt-Jakob disease (CJD), the possible occurrence of variant CJD (vCJD), and potential zoonotic transmission of chronic wasting disease (CWD). Although cross- species transmission of prion diseases seems to be limited by an apparent “species barrier,” the occurrence of bovine spongiform encephalopathy (BSE) and its transmis- sion to humans indicate that animal prion diseases can pose a significant public health risk. Recent reports of secondary person-to-person spread of vCJD via blood products and detection of vCJD transmission in a patient heterozygous at codon 129 further illustrate the potential public health impacts of BSE. INTRODUCTION by IRMO/Information Center on 03/14/05. For personal use only. Prion diseases, also known as transmissible spongiform encephalopathies (TSEs), are a group of animal and human brain diseases that are uniformly fatal and often characterized by a long incubation period and a multifocal neuropathologic picture of neuronal loss, spongiform changes, and astrogliosis (3). -

Possible Modes of Transmission of Novel Coronavirus SARS-Cov-2
Acta Biomed 2020; Vol. 91, N. 3: e2020036 DOI: 10.23750/abm.v91i3.10039 © Mattioli 1885 Reviews / Focus on Possible modes of transmission of Novel Coronavirus SARS-CoV-2: a review Richa Mukhra1, Kewal Krishan1, Tanuj Kanchan2 1 Department of Anthropology (UGC Centre of Advanced Study), Panjab University, Sector-14, Chandigarh, India 2 Department of Forensic Medicine and Toxicology, All India Institute of Medical Sciences, Jodhpur, India. Abstract: Introduction: The widespread outbreak of the novel SARS-CoV-2 has raised numerous questions about the origin and transmission of the virus. Knowledge about the mode of transmission as well as assess- ing the effectiveness of the preventive measures would aid in containing the outbreak of the coronavirus. Presently, respiratory droplets, physical contact and aerosols/air-borne have been reported as the modes of SARS-CoV-2 transmission of the virus. Besides, some of the other possible modes of transmission are being explored by the researchers, with some studies suggesting the viral spread through fecal-oral, conjunctival secretions, flatulence (farts), sexual and vertical transmission from mother to the fetus, and through asymp- tomatic carriers, etc. Aim: The primary objective was to review the present understanding and knowledge about the transmission of SARS-CoV-2 and also to suggest recommendations in containing and preventing the novel coronavirus. Methods: A review of possible modes of transmission of the novel SARS-CoV-2 was conducted based on the reports and articles available in PubMed and ScienceDirect.com that were searched using keywords, ‘transmission’, ‘modes of transmission’, ‘SARS-CoV-2’, ‘novel coronavirus’, and ‘COVID-19’. Articles refer- ring to air-borne, conjunctiva, fecal-oral, maternal-fetal, flatulence (farts), and breast milk transmission were included, while the remaining were excluded. -

Innate Immunity Part I: the Immediate Response Deborah A
Innate Immunity Part I: The Immediate Response Deborah A. Lebman, Ph.D. OBJECTIVES 1. Describe three mechanisms present at epithelial surfaces that protect the host from pathogens. 2. List 3 functions of complement proteins in innate immunity 3. Know the primary functions of macrophages and neutrophils 4. List the receptors on macrophages involved in recognition of pathogens 5. List the key toxic products of phagocytes READING • Parham, The Immune System, pg 227-238. TERMS • Defensin: antibacterial peptide produced by the host at epithelial surfaces • Complement: ubiquitously present proteins synthesized primarily in the liver that mediate a variety of responses to pathogens • Opsonin (opsonization): alteration of cell surface that facilitates phagocytosis • Phagocytosis: internalization of particulate matter by cells • CD14: LPS receptor • TLR-4: toll like receptor 4 I. Introduction A. The first response to pathogens occurs immediately B. The early response employs both mechanical and cell mediated mechanisms C. There is no cellular expansion, but there is cell recruitment D. A wide array of soluble mediators with both positive and negative effects, i.e. remove the pathogen but hurt the host, is released II. Overview of Pathogens A. There are four types of pathogen: bacteria, virus, fungus, parasite B. Table I: Examples of the Four Types of Common Human Pathogen Table 1: Examples of the Four Types of Common Human Pathogen Table 1 (Continued): Examples of the Four Types of Common Human Pathogen C. Pathogens can be intracellular or extracellular Table 2: Examples of intracellular and extracellular pathogens. Notice that both innate and adaptive mechanisms are involved in the response to both types of pathogen. -

Intersecting Infections of Public Health Significance Page I Intersecting Infections of Public Health Significance
Intersecting Infections of Public Health Significance Page i Intersecting Infections of Public Health Significance The Epidemiology of HIV, Viral Hepatitis, Sexually Transmitted Diseases, and Tuberculosis in King County 2008 Intersecting Infections of Public Health Significance was supported by a cooperative agreement from the Centers for Disease Control and Prevention Published December 2009 Alternate formats of this report are available upon request Intersecting Infections of Public Health Significance Page ii David Fleming, MD, Director Jeffrey Duchin, MD, Director, Communicable Disease Epidemiology & Immunization Program Matthew Golden, MD, MPH, Director, STD Program Masa Narita, MD, Director, TB Program Robert Wood, MD, Director, HIV/AIDS Program Prepared by: Hanne Thiede, DVM, MPH Elizabeth Barash, MPH Jim Kent, MS Jane Koehler, DVM, MPH Other contributors: Amy Bennett, MPH Richard Burt, PhD Susan Buskin, PhD, MPH Christina Thibault, MPH Roxanne Pieper Kerani, PhD Eyal Oren, MS Shelly McKeirnan, RN, MPH Amy Laurent, MSPH Cover design and formatting by Tanya Hunnell The report is available at www.kingcounty.gov/health/hiv For additional copies of this report contact: HIV/AIDS Epidemiology Program Public Health – Seattle & King County 400 Yesler Way, 3rd Floor Seattle, WA 98104 206-296-4645 Intersecting Infections of Public Health Significance Page iii Table of Contents INDEX OF TABLES AND FIGURES ········································································ vi EXECUTIVE SUMMARY ······················································································ -
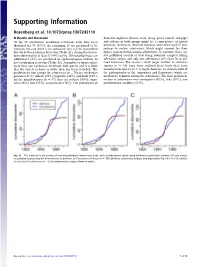
Supporting Information
Supporting Information Rosenberg et al. 10.1073/pnas.1307243110 SI Results and Discussion domestic ungulates (horses, cows, sheep, goats, camels, and pigs) Of the 83 arboviruses, nonhuman vertebrate hosts have been and rodents in both groups might be a consequence of spatial identified for 70 (84%); the remaining 13 are presumed to be proximity to humans. Sentinel monkeys were often used in pro- zoonoses because there is no indication they can be transmitted cedures to isolate arboviruses, which might account for their directly between humans by vectors (Table S1). Animal hosts have higher representation among arboviruses. In contrast, there are been identified for at least 57 (44%) of the 130 nonarboviruses; an few published records of bats being routinely sampled during additional 5 (8%) are presumed on epidemiological evidence to arbovirus studies, and only two arboviruses (3%) have been iso- have nonhuman reservoirs (Table S1). A number of viruses infect lated from bats. The reason a much larger number of arbovirus more than one nonhuman vertebrate host species and it is likely species (n = 16) have been isolated from birds than have that the variety of hosts is wider than has been recorded. The nonarbovirus species (n = 1) might, however, be characteristic of predominant host groups for arboviruses (n = 70) are nonhuman the pathogenicity of the togaviruses and flaviviruses, which are primates (31%), rodents (29%), ungulates (26%), and birds (23%); much more common among the arboviruses. The most prominent for the nonarboviruses (n = 57), they are rodents (30%), ungu- vectors of arboviruses were mosquitoes (67%), ticks (19%), and lates (26%), bats (23%), and primates (16%). -

Spillback in the Anthropocene: the Risk of Human-To-Wildlife Pathogen Transmission for Conservation and Public Health
Spillback in the Anthropocene: the risk of human-to-wildlife pathogen transmission for conservation and public health Anna C. Fagre*1, Lily E. Cohen2, Evan A. Eskew3, Max Farrell4, Emma Glennon5, Maxwell B. Joseph6, Hannah K. Frank7, Sadie Ryan8, 9,10, Colin J Carlson11,12, Gregory F Albery*13 *Corresponding authors: [email protected]; [email protected] 1: Department of Microbiology, Immunology, and Pathology, College of Veterinary Medicine and Biomedical Sciences, Colorado State University, Fort Collins, CO 2: Icahn School of Medicine at Mount Sinai, New York, NY 10029 3: Department of Biology, Pacific Lutheran University, Tacoma, WA, 98447 USA 4: Department of Ecology & Evolutionary Biology, University of Toronto 5: Disease Dynamics Unit, Department of Veterinary Medicine, University of Cambridge, Cambridge CB3 0ES, UK 6: Earth Lab, University of Colorado Boulder, Boulder, CO 80309 7: Department of Ecology and Evolutionary Biology, Tulane University, New Orleans, LA, 70118 USA 8: Quantitative Disease Ecology and Conservation (QDEC) Lab Group, Department of Geography, University of Florida, Gainesville, FL, 32610 USA 9: Emerging Pathogens Institute, University of Florida, Gainesville, FL, 32610 USA 10: School of Life Sciences, University of KwaZulu-Natal, Durban, 4041, South Africa 11: Center for Global Health Science and Security, Georgetown University Medical Center, Washington, DC, 20057 USA 12: Department of Microbiology and Immunology, Georgetown University Medical Center, Washington, DC, 20057 USA 13: Department of Biology, Georgetown University, Washington, DC, 20057 USA 1 Abstract The SARS-CoV-2 pandemic has led to increased concern over transmission of pathogens from humans to animals (“spillback”) and its potential to threaten conservation and public health.