A Taxonomic Revision of Rhododendron L. Section Pentanthera G
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Phylogenetics, Flow-Cytometry and Pollen Storage in Erica L
Institut für Nutzpflanzenwissenschaft und Res sourcenschutz Professur für Pflanzenzüchtung Prof. Dr. J. Léon Phylogenetics, flow-cytometry and pollen storage in Erica L. (Ericaceae). Implications for plant breeding and interspecific crosses. Inaugural-Dissertation zur Erlangung des Grades Doktor der Agrarwissenschaften (Dr. agr.) der Landwirtschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität Bonn von Ana Laura Mugrabi de Kuppler aus Buenos Aires Institut für Nutzpflanzenwissenschaft und Res sourcenschutz Professur für Pflanzenzüchtung Prof. Dr. J. Léon Referent: Prof. Dr. Jens Léon Korreferent: Prof. Dr. Jaime Fagúndez Korreferent: Prof. Dr. Dietmar Quandt Tag der mündlichen Prüfung: 15.11.2013 Erscheinungsjahr: 2013 A mis flores Rolf y Florian Abstract Abstract With over 840 species Erica L. is one of the largest genera of the Ericaceae, comprising woody perennial plants that occur from Scandinavia to South Africa. According to previous studies, the northern species, present in Europe and the Mediterranean, form a paraphyletic, basal clade, and the southern species, present in South Africa, form a robust monophyletic group. In this work a molecular phylogenetic analysis from European and from Central and South African Erica species was performed using the chloroplast regions: trnL-trnL-trnF and 5´trnK-matK , as well as the nuclear DNA marker ITS, in order i) to state the monophyly of the northern and southern species, ii) to determine the phylogenetic relationships between the species and contrasting them with previous systematic research studies and iii) to compare the results provided from nuclear data and explore possible evolutionary patterns. All species were monophyletic except for the widely spread E. arborea , and E. manipuliflora . The paraphyly of the northern species was also confirmed, but three taxa from Central East Africa were polyphyletic, suggesting different episodes of colonization of this area. -

A STUDY of the TYRONE - MOUNT Udion LINEAMENT by REMOTE SENSING TECHNIWES and Fleld NIETHQDS
A STUDY OF THE TYRONE - MOUNT UdION LINEAMENT BY REMOTE SENSING TECHNIWES AND FlELD NIETHQDS Con- NAS 5-22822 0. P. Gold, Principal Indga@x ORSER Technical Report 12-77 Office for Ramow Sensing of Earth Resources 219 Electrical Engineering West University Park, PA 16802 Prepared for GODDARD SPACE FLIGHT CENTER Greenbelt, MD 20771 The field invesrigations and much of the analvsis descrihrd in this report were used as the basis for a payer in Crology. submtttrd by Elicharl R. Cnnich to tbe Departmc~~tof ~:t~oscic-~\c~?i, in partial iu!ffllment of the rrqufrrurznts tor tlrc Mastt-r- oi Science Dcgrre. b I r T-nd- r RoonOlk I A S~~M)YOF ms ~RONR- ~\nurWIW LIN~BY December 1977 RBWTS SMSSIUC TECHNIQUS AND FlliLD t4STHWS 6 hnwym-- - r Technical Report 12-?7 J 7. habftd a -@-mQImwwn-amL David P. Gold, Principal Inwe8tigator m , 10. WRk uau Wa e m)a~~mt-#y#mdMem Office for Remote Senaing of Earth Resources 219 Electrical Bngineerin8 West Building rt. bn~nar ~nn\ &. \ The Pennsylvania State Uniwrsi ty NA!! 5-22822 University Park, PA 16802 13 ~*oro@~.om.ndhd~CI tr sponss~lr-v-nd~ddnr Final Report 11 1176-6/30/77 Caddard Space Flight Center I Crecnbelt, ND 20771 4 m.LI Field work was combined with satellite imagery and photography to study the Tyrone - Pkrunt Union lineament in Blair and Huntingdon Counties, central Pennsylvania. This feature, expressed as the valleys containing the Little Juniata and Juniata River: . transgtesscs the nose of the southwest-plunging Nittany Anticlinorium in the western extremity of the Valley ;~ndRidge Province. -
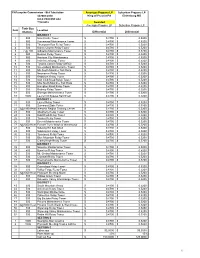
BID TAB Propane 2018-2023.Xlsx
PATurnpike Commission - Bid Tabulation Amerigas Propane LP Suburban Propane LP 447002-2018 King of Prussia PA Eldersburg MD BULK PROPANE GAS 7/12/2018 Awarded Amerigas Propane LP Suburban Propane LP Tank Size Item Location (Gallons) Differential Differential DISTRICT 1 1 500 New Castle Tower$ 0.4790 $ 0.5200 2 500 Homewood Maintenance Tower $ 0.4790 $ 0.5200 3 500 Thompson Run Relay Tower$ 0.4790 $ 0.5200 4 500 Salem Church Relay Tower$ 0.4790 $ 0.5200 5 2 @ 100 Gibsonia Maintenance Tower $ 0.4790 $ 0.5200 6 500 Beacon Relay Tower$ 0.4790 $ 0.5200 7 500 Harrison City Maintenance $ 0.4790 $ 0.5200 8 500 Irwin Interchange Tower$ 0.4790 $ 0.5200 9 500 Tyson's Corner Relay (WRO)$ 0.4790 $ 0.5200 10 500 Greensburg Maintenance Tower$ 0.4790 $ 0.5200 11 500 M5 South Mainline Toll Plaza$ 0.4790 $ 0.5200 12 500 Newcomer Relay Tower$ 0.4790 $ 0.5200 13 500 Redstone Relay Tower$ 0.4790 $ 0.5200 14 500 Gun Club Road Relay Tower$ 0.4790 $ 0.5200 15 500 M52 North Mainline Toll Plaza$ 0.4790 $ 0.5200 16 500 Hampton Road Relay Tower $ 0.4790 $ 0.5200 17 500 Rodney Relay Tower$ 0.4790 $ 0.5200 18 500 Donegal Maintenenace Tower $ 0.4790 $ 0.5200 19 1000 Laurel Hill Bypass Salt Shed$ 0.4790 $ 0.5200 DISTRICT 2 20 500 Laurel Relay Tower$ 0.4790 $ 0.5200 21 500 Somerset State Police $ 0.4790 $ 0.5200 22 2@1990 Heat Western Region Training Center$ 0.4790 $ 0.5200 23 500 Allegheny Relay Tower$ 0.4790 $ 0.5200 24 500 Bald Knob Relay Tower$ 0.4790 $ 0.5200 25 500 Tussey Relay Tower$ 0.4790 $ 0.5200 26 500 Everett Maintenance Tower$ 0.4790 $ 0.5200 27 2@1000 Heat Everett Truck Wash & Warehouse$ 0.4790 $ 0.5200 28 500 Sideling Hill Salt Shed $ 0.4790 $ 0.5200 29 500 Sideling Hill Relay Tower$ 0.4790 $ 0.5200 30 500 Tuscarora Relay Tower$ 0.4790 $ 0.5200 31 500 Blue Mountain Relay Tower$ 0.4790 $ 0.5200 32 500 Clarks Knob Relay Tower$ 0.4790 $ 0.5200 DISTRICT 3 33 500 Newville Maintenance Tower$ 0.4790 $ 0.5200 34 500 Bunches Relay Tower$ 0.4790 $ 0.5200 35 500 Mt. -
1835. EXECUTIVE. *L POST OFFICE DEPARTMENT
1835. EXECUTIVE. *l POST OFFICE DEPARTMENT. Persons employed in the General Post Office, with the annual compensation of each. Where Compen Names. Offices. Born. sation. Dol. cts. Amos Kendall..., Postmaster General.... Mass. 6000 00 Charles K. Gardner Ass't P. M. Gen. 1st Div. N. Jersey250 0 00 SelahR. Hobbie.. Ass't P. M. Gen. 2d Div. N. York. 2500 00 P. S. Loughborough Chief Clerk Kentucky 1700 00 Robert Johnson. ., Accountant, 3d Division Penn 1400 00 CLERKS. Thomas B. Dyer... Principal Book Keeper Maryland 1400 00 Joseph W. Hand... Solicitor Conn 1400 00 John Suter Principal Pay Clerk. Maryland 1400 00 John McLeod Register's Office Scotland. 1200 00 William G. Eliot.. .Chie f Examiner Mass 1200 00 Michael T. Simpson Sup't Dead Letter OfficePen n 1200 00 David Saunders Chief Register Virginia.. 1200 00 Arthur Nelson Principal Clerk, N. Div.Marylan d 1200 00 Richard Dement Second Book Keeper.. do.. 1200 00 Josiah F.Caldwell.. Register's Office N. Jersey 1200 00 George L. Douglass Principal Clerk, S. Div.Kentucky -1200 00 Nicholas Tastet Bank Accountant Spain. 1200 00 Thomas Arbuckle.. Register's Office Ireland 1100 00 Samuel Fitzhugh.., do Maryland 1000 00 Wm. C,Lipscomb. do : for) Virginia. 1000 00 Thos. B. Addison. f Record Clerk con-> Maryland 1000 00 < routes and v....) Matthias Ross f. tracts, N. Div, N. Jersey1000 00 David Koones Dead Letter Office Maryland 1000 00 Presley Simpson... Examiner's Office Virginia- 1000 00 Grafton D. Hanson. Solicitor's Office.. Maryland 1000 00 Walter D. Addison. Recorder, Div. of Acc'ts do.. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Learn About Plant Diversity, with Focus on the Angiosperm Flora of Washington State
Biol 317: Plant Classification & Identification Spring 2014 Instructor: Dr. Pat Lu-Irving Office: 408 Hitchcock Office hours: Tue & Wed, 10-11am (or by appointment) Email: [email protected] Grad TA: John Chau Peer TAs: Adrienne Cohen Sam Frankel Kate Nowell Veanna Willard Class website: http://courses.washington.edu/bot113/ Course objectives Learn about plant diversity, with focus on the angiosperm flora of Washington state. Most land plants are flowering plants Incl. grasses, roses, lilies, daisies; producing both flowers and fruits WA natives There are approximately 300,000 species of angiosperms, with 3,000 occurring in WA Compare with mammals: only 5,500 on Earth Course objectives Understand the principles and philosophy of plant classification. In modern classification, our goal is to recognize monophyletic groups Monocots Eudicots Knowledge of phylogeny is vital to plant classification Course objectives Identify 40 important plant families by sight. Key unknown plants to species using published floras. Orchidaceae Araceae Knowledge of morphology is vital to plant identification sepal corolla Corallorhiza maculata Lysichiton americanus stem Ericaceae Boraginaceae stamen leaf ovary Elliottia pyroliflora Myosotis sylvatica Course objectives Gain a greater appreciation for nature, and your connection Triticum aestivum with it. Hevea brasiliensis Wheat flour Natural rubber Coffea arabica Erythroxylum coca Coffee Course objectives Gain a greater appreciation for nature, and your connection with it. Today’s lecture Nomenclature and classification -

Pennsylvania
Elton 243 13A Road Ithaca Harford 36 54A 226 220 Dayton 390 14 79 11 41 80° 30’ 80° 15’ 80° 00’ 79° 45’ 79° 30’ Toll 79° 15’ 79° 00’ 78° 45’ 78° 30’Machias 78° 15’ 78° 00’ 77° 45’ 77° 30’ 77° 15’ 77° 00’ 76° 45’ 76° 30’ 76° 15’ 76° 00’ 75° 45’ 75° 30’ 75° 15’ 75° 00’ 74° 45’ 74° 30’ Houghton 53 414 5 38 21 Haskinville 327 26 A B C D E F G H J K 3 L M N 1 O P Q R S T U V W X Y Z To Buffalo East Otto Rushford Arkport Avoca 36 2 54 Hammondsport Caroline 219 Caneadea 3 96B Whitney 60 83 Hamlet South 62 243 Watkins 10 20 Cassadaga 240 98 Birdsall 10 415 37 13 79 Point 7 90 Dayton 16 2 86 35 7 Glen 3 Montour 228 THESE Bloomville Bear 322 5 Wesley Lake 41 12 TOURISM AND TRANSPORTATION28 MAP Lakes 353 242 17 79 Barcelona Lake Belfast 3 Falls Danby Center Lisle Triangle 206 Unadilla Franklin Cassadaga 6 34 329 Newfield Lisle 60 Howard Richford 11 357 Tom Wolf, Governor 1 1 2 19 Bath Townsend 224 QUICK RESPONSE 8 Greene CoventryPA Sidney PA 5 5 3 33 Hornell 38 Sonora 29 34 2 Whitney Point 41 10 Westfield Charlotte Cattaraugus Franklinville Odessa 206 Stockton 9 96 Yassmin Gramian, P.E., Secretary, Department of Transportation Center Cherry Ashford Almond 4 30 9 Creek Angelica 32 Speedsville Itaska visitPA.com41 Bainbridge visitPA.com STATE 17 7 226 Monterey 414 CODES N 394 Leon 8 42 39 5 8 88 9 Delhi Forsyth 21 86 Willseyville ROAD CLASSIFICATION AND ROUTE MARKERS KEY TO MAP SYMBOLS 61 6 New 36 3 Beaver Dams 7 20 PARKSAlbion 5 Nanticoke TRAFFIC INTERCHANGES 28 ? Hartfield 27 353 31 Alfred Savona 16 CAN BE USEDJenksville Glen 7 PASSENGER RAILROADS -
View of Valley and Ridge Structures from ?:R Stop IX
GIJIDEBOOJ< TECTONICS AND. CAMBRIAN·ORDO'IICIAN STRATIGRAPHY CENTRAL APPALACHIANS OF PENNSYLVANIA. Pifftbutgh Geological Society with the Appalachian Geological Society Septembet, 1963 TECTONICS AND CAMBRIAN -ORDOVICIAN STRATIGRAPHY in the CENTRAL APPALACHIANS OF PENNSYLVANIA FIELD CONFERENCE SPONSORS Pittsburgh Geological Society Appalachian Geological Society September 19, 20, 21, 1963 CONTENTS Page Introduction 1 Acknowledgments 2 Cambro-Ordovician Stratigraphy of Central and South-Central 3 Pennsylvania by W. R. Wagner Fold Patterns and Continuous Deformation Mechanisms of the 13 Central Pennsylvania Folded Appalachians by R. P. Nickelsen Road Log 1st day: Bedford to State College 31 2nd day: State College to Hagerstown 65 3rd day: Hagerstown to Bedford 11.5 ILLUSTRATIONS Page Wagner paper: Figure 1. Stratigraphic cross-section of Upper-Cambrian 4 in central and south-central Pennsylvania Figure 2. Stratigraphic section of St.Paul-Beekmantown 6 rocks in central Pennsylvania and nearby Maryland Nickelsen paper: Figure 1. Geologic map of Pennsylvania 15 Figure 2. Structural lithic units and Size-Orders of folds 18 in central Pennsylvania Figure 3. Camera lucida sketches of cleavage and folds 23 Figure 4. Schematic drawing of rotational movements in 27 flexure folds Road Log: Figure 1. Route of Field Trip 30 Figure 2. Stratigraphic column for route of Field Trip 34 Figure 3. Cross-section of Martin, Miller and Rankey wells- 41 Stops I and II Figure 4. Map and cross-sections in sinking Valley area- 55 Stop III Figure 5. Panorama view of Valley and Ridge structures from ?:r Stop IX Figure 6. Camera lucida sketch of sedimentary features in ?6 contorted shale - Stop X Figure 7- Cleavage and bedding relationship at Stop XI ?9 Figure 8. -

Alkaline Addition Problems at the Skytop/Interstate-99 Site, Central Pennsylvania1
ALKALINE ADDITION PROBLEMS AT THE SKYTOP/INTERSTATE-99 SITE, CENTRAL PENNSYLVANIA1 Arthur W. Rose and Hubert L. Barnes2 Abstract. In 2002-3, approximately 8x105 m3 of pyrite-bearing Bald Eagle sandstone and Reedsville shale were removed from a large road cut through Bald Eagle Ridge near State College, PA. The rock contained an average of about 4.5% pyrite as veins and fine veinlet networks across 200 m of road cut. Smaller amounts of ZnS and other heavy metal sulfides accompany the pyrite. The rock was placed in 9 nearby locations, including large waste piles and several valley fills, two ‘buttresses” and a lane elevation along about 0.8 km of hillside that was threatening to slide into the road. During excavation of the cut, pyrite was recognized as a potential problem and considerable lime was added as layers to the various piles. Despite the lime addition, highly acidic seeps emerged from the piles and fills, with pH 2.0 to 2.7, Fe 90-1500 mg/L, and acidities as high as 18,000 mg/L (as CaCO3). These results clearly show that addition of lime and other alkaline materials as layers is not effective. In experiments to test remediation methods, Bauxsol slurry was sprayed onto part of the buttress area but failed to prevent continuing acid seepages. Inspection trenches showed little penetration of the Bauxsol, and demonstrated the presence of the added lime as impermeable lime layers within the buttress. Bucket tests of mixtures of alkaline circulating fluidized-bed ash with pyritic rocks, when well mixed, gave alkaline effluents. -

The Civilian Conservation Corps and the National Park Service, 1933-1942: an Administrative History. INSTITUTION National Park Service (Dept
DOCUMENT RESUME ED 266 012 SE 046 389 AUTHOR Paige, John C. TITLE The Civilian Conservation Corps and the National Park Service, 1933-1942: An Administrative History. INSTITUTION National Park Service (Dept. of Interior), Washington, D.C. REPORT NO NPS-D-189 PUB DATE 85 NOTE 293p.; Photographs may not reproduce well. PUB TYPE Reports - Descriptive (141) -- Historical Materials (060) EDRS PRICE MF01/PC12 Plus Postage. DESCRIPTORS *Conservation (Environment); Employment Programs; *Environmental Education; *Federal Programs; Forestry; Natural Resources; Parks; *Physical Environment; *Resident Camp Programs; Soil Conservation IDENTIFIERS *Civilian Conservation Corps; Environmental Management; *National Park Service ABSTRACT The Civilian Conservation Corps (CCC) has been credited as one of Franklin D. Roosevelt's most successful effortsto conserve both the natural and human resources of the nation. This publication provides a review of the program and its impacton resource conservation, environmental management, and education. Chapters give accounts of: (1) the history of the CCC (tracing its origins, establishment, and termination); (2) the National Park Service role (explaining national and state parkprograms and co-operative planning elements); (3) National Park Servicecamps (describing programs and personnel training and education); (4) contributions of the CCC (identifying the major benefits ofthe program in the areas of resource conservation, park and recreational development, and natural and archaeological history finds); and (5) overall -

Curt Teich Postcard Archives Towns and Cities
Curt Teich Postcard Archives Towns and Cities Alaska Aialik Bay Alaska Highway Alcan Highway Anchorage Arctic Auk Lake Cape Prince of Wales Castle Rock Chilkoot Pass Columbia Glacier Cook Inlet Copper River Cordova Curry Dawson Denali Denali National Park Eagle Fairbanks Five Finger Rapids Gastineau Channel Glacier Bay Glenn Highway Haines Harding Gateway Homer Hoonah Hurricane Gulch Inland Passage Inside Passage Isabel Pass Juneau Katmai National Monument Kenai Kenai Lake Kenai Peninsula Kenai River Kechikan Ketchikan Creek Kodiak Kodiak Island Kotzebue Lake Atlin Lake Bennett Latouche Lynn Canal Matanuska Valley McKinley Park Mendenhall Glacier Miles Canyon Montgomery Mount Blackburn Mount Dewey Mount McKinley Mount McKinley Park Mount O’Neal Mount Sanford Muir Glacier Nome North Slope Noyes Island Nushagak Opelika Palmer Petersburg Pribilof Island Resurrection Bay Richardson Highway Rocy Point St. Michael Sawtooth Mountain Sentinal Island Seward Sitka Sitka National Park Skagway Southeastern Alaska Stikine Rier Sulzer Summit Swift Current Taku Glacier Taku Inlet Taku Lodge Tanana Tanana River Tok Tunnel Mountain Valdez White Pass Whitehorse Wrangell Wrangell Narrow Yukon Yukon River General Views—no specific location Alabama Albany Albertville Alexander City Andalusia Anniston Ashford Athens Attalla Auburn Batesville Bessemer Birmingham Blue Lake Blue Springs Boaz Bobler’s Creek Boyles Brewton Bridgeport Camden Camp Hill Camp Rucker Carbon Hill Castleberry Centerville Centre Chapman Chattahoochee Valley Cheaha State Park Choctaw County -

Montana Map List
Montana Map List Maps may be ordered online at http://store.usgs.gov For more information, call 1-888-ASK-USGS Distributed courtesy www.railroads-of-montana.com UNITED STATES GEOLOGICAL SURVEY USGS Quadrangle Name Quadrangle Number Version Year Scale Latitude Coordinates Longitude Coordinates ABBOTT LAKE, MT 9780607321890 01/01/1985 1:24,000 482230 - 483000 -1121500 - -1120730 ABSAROKEE, MT 9780607295627 01/01/1955 1:24,000 453000 - 453730 -1093000 - -1092230 ACKLEY LAKE, MT 9780607295634 01/01/1970 1:24,000 465230 - 470000 -1100000 - -1095230 ACME, WY 9780607600667 01/01/1978 1:24,000 445230 - 450000 -1070000 - -1065230 ACORN FLATS, MT 9780607295641 01/01/1965 1:24,000 464500 - 465230 -1070730 - -1070000 ACTON, MT 9780607295658 01/01/1956 1:24,000 455230 - 460000 -1084500 - -1083730 ADA CREEK, MT 9780607295665 01/01/1964 1:24,000 471500 - 472230 -1063730 - -1063000 ADAIR, ID 9780607891607 01/01/1995 1:24,000 471500 - 472230 -1153730 - -1153000 AGAWAM NORTH, MT 9780607321906 01/01/1985 1:24,000 480000 - 480730 -1121500 - -1120730 AGAWAM SOUTH, MT 9780607322538 01/01/1987 1:24,000 475230 - 480000 -1121500 - -1120730 AHERN PASS, MT 9780607295672 01/01/1968 1:24,000 484500 - 485230 -1135230 - -1134500 AHLES, MT 9780607295689 01/01/1983 1:24,000 462230 - 463000 -1071500 - -1070730 AJAX RANCH, MT 9780607996111 01/01/1997 1:24,000 452230 - 453000 -1133730 - -1133000 AKERS RANCH, MT 9780607295702 01/01/1963 1:24,000 463730 - 464500 -1080730 - -1080000 ALASKA BENCH, MT 9780607880083 01/01/1995 1:24,000 464500 - 465230 -1091500 - -1090730 ALBERTON,