Integrated Biostratigraphy Across the Aalenian/Bajocian Boundary of the Central High Atlas, Morocco
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Jurassic Pleurotomarioidean Gastropod Laevitomaria and Its Palaeobiogeographical History
The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history ROBERTO GATTO, STEFANO MONARI, JÁNOS SZABÓ, and MARIA ALESSANDRA CONTI Gatto, R., Monari, S., Szabó, J., and Conti, M.A. 2015. The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history. Acta Palaeontologica Polonica 60 (1): 217–233. The genus Laevitomaria is reviewed and its palaeobiogeographical history is reconstructed based on the re-examination of its type species L. problematica, the study of material stored at the National Natural History Museum of Luxembourg, and an extensive review of the literature. The systematic study allows ascribing to Laevitomaria a number of Jurassic species from the western European region formerly included in other pleurotomariid genera. The following new combi- nations are proposed: Laevitomaria allionta, L. amyntas, L. angulba, L. asurai, L. daityai, L. fasciata, L. gyroplata, L. isarensis, L. joannis, L. repeliniana, L. stoddarti, L. subplatyspira, and L. zonata. The genus, which was once considered as endemic of the central part of the western Tethys, shows an evolutionary and palaeogeographical history consider- ably more complex than previously assumed. It first appeared in the Late Sinemurian in the northern belt of the central western Tethys involved in the Neotethyan rifting, where it experienced a first radiation followed by an abrupt decline of diversity in the Toarcian. Species diversity increased again during Toarcian–Aalenian times in the southernmost part of western European shelf and a major radiation occurred during the Middle Aalenian to Early Bajocian in the northern Paris Basin and southern England. After a latest Bajocian collapse of diversity, Laevitomaria disappeared from both the central part of western Tethys and the European shelf. -
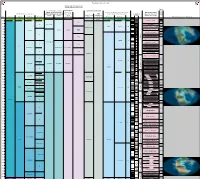
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

A Key Interval in the Early Mesozoic Phytoplankton Radiation
1 The Bajocian (Middle Jurassic): a key interval in the early Mesozoic phytoplankton radiation 2 3 Nickolas J. Wiggan a.b*, James B. Riding b, Robert A. Fensome c, Emanuela Mattioli d 4 5 Insert addresses here 6 7 ABSTRACT 8 Dinoflagellates and coccolithophores are two of the most important groups of phytoplankton 9 in the modern oceans. These groups originated in the Triassic and radiated through the early 10 Mesozoic, rising to ecological prominence. Within this long-term radiation, important short- 11 term intervals of evolutionary and ecological change can be recognised. The Bajocian 12 (Middle Jurassic, ~170–168 Ma) was characterised by an important ecological transition 13 within the coccolithophores, and the radiation of one of the principal families of cyst-forming 14 dinoflagellates, the Gonyaulacaceae. During the Early Bajocian, the coccolith genus 15 Watznaueria diversified and expanded ecologically to dominate coccolith floras, a situation 16 which continued for the remainder of the Mesozoic. This pattern was paralleled within 17 dinoflagellate cyst floras by the ecological dominance of the genus Dissiliodinium in the mid- 18 palaeolatitudes. These phenomena appear to be linked to a positive carbon isotope shift, and 19 an interval of enhanced productivity driven by a shift to a more humid climate, enhanced 20 continental weathering and nutrient flux. The latest Early Bajocian to earliest Bathonian was 21 then characterised by the rapid increase in diversity of dinoflagellate cysts within the family 22 Gonyaulacaceae. Through this interval, the Gonyaulacaceae transitioned from being a 23 relatively minor component of dinoflagellate cyst floras, to becoming one of the prominent 24 groups of cyst-forming dinoflagellates, which has persisted to the Holocene. -

Paléontologie Au Luxembourg (2) A
Paléontologie au Luxembourg (2) A. Di Cencio, D. Sadki, R. Weis (eds.) R. Weis D. Sadki, A. Di Cencio, Les ammonites de la Minette Andrea Di Cencio, Driss Sadki, Paléontologie au Luxembourg (2) Luxembourg au Paléontologie Robert Weis (eds.) Ferrantia Travaux scientifiques du Musée national d'histoire naturelle Luxembourg www.mnhn.lu 83 2020 Ferrantia 83 2020 2020 83 Ferrantia est une revue publiée à intervalles non réguliers par le Musée national d’histoire naturelle à Luxembourg. Elle fait suite, avec la même tomaison, aux T M ’ L parus entre 1981 et 1999. Comité de rédaction: Eric Buttini Guy Colling Alain Frantz Thierry Helminger Ben Thuy Mise en page: Romain Bei Design: Thierry Helminger Prix du volume: 20 € Rédaction: Échange: Musée national d’histoire naturelle Exchange MnhnL Rédaction Ferrantia c/o Musée national d’histoire naturelle 25, rue Münster 25, rue Münster L-2160 Luxembourg L-2160 Luxembourg Tél +352 46 22 33 - 1 Tél +352 46 22 33 - 1 Fax +352 46 38 48 Fax +352 46 38 48 Internet: http://www.mnhn.lu/ferrantia/ Internet: http://www.mnhnl.lu/biblio/exchange email: [email protected] email: [email protected] Page de couverture: Bredyia subinsignis (Oppel, 1856), DOU833. Natural History Museum of Luxembourg. Citation: Di Cencio A., Sadki D., Weis R. (eds.) 2020. - Paléontologie au Luxembourg (2) - Les ammonites de la Minette. Ferrantia 83, Musée national d’histoire naturelle, Luxembourg, 129 p. Date de publication: 15 décembre 2020 (réception du manuscrit: 19 mai 2020) Impression: Imprimerie Centrale climatiquement neutre Impression | LU-319-JR8FDJV | www.natureOffice.com Ferrantia est publiée sous la licence Creative Commons BY-NC-ND 3.0 LU. -

The Graphoceratid Ammonite Succession in the Aalenian and Lowest Bajocian (Middle Jurassic) at Horn Park, Dorset, UK ROBERT B
The Graphoceratid Ammonite Succession in the Aalenian and lowest Bajocian (Middle Jurassic) at Horn Park, Dorset, UK ROBERT B. CHANDLER Riddlesdown High School, Purley, Surrey, CR8 lEX Summary In 1990 Callomon and Chandler set out a scheme of 16 faunal horizons for the Aalenian-Lower Bajocian of Dorset and Somerset based principally on graphoceratid ammonites. One locality, Horn Park, stands apart in displaying strata from almost all the horizons identified in that work. Yet little material from it has ever been figured. Here a section is given together with exhaustive tables of existing nominal morphospecies collected at each level and with figures that allow the variability of the suc- cessive, probably in most cases, monobiospecific assemblages to be assessed. A revised generic classification of the Graphoceratidae is presented. them precludes almost any satisfactory discussion of phyletic relationships and hence of the overall evolution The highly discontinuous, condensed and rapidly of the group. varying development of the Inferior Oolite in southern England was first perceived by S.S. Buckman (e.g. 1891, The biostratigraphy of the Graphoceratidae has now 1893, 1910) during his attempts to correlate sections by been rather well documented, for example by Rocha et means of ammonites. Leading amongst these in al. (1990), Henriques (1992; et al. 1994) in Portugal; the Aalenian part of the succession are the Linares and Sandoval (1990) in Spain, Sadki (1994) in Graphoceratidae, including such well known genera as Morocco. In Britain, Morton (1990; 1994) and Morton Leioceras, Ludwigia, Brasilia, Graphoceras and and Hudson (1995) have described the succession in Hyperlioceras. Scotland and compared it with Dorset (Morton and Chandler 1994). -

Jurassic (170 - 199 Ma Time-Slice) Time
Early Jurassic (170 - 199 Ma time-slice) Time ScaLe R Creator CHRONOS Cen Mesozoic Updated by James G. Ogg (Purdue University) and Gabi Ogg to: GEOLOGIC TIME SCALE 2004 (Gradstein, F.M., Ogg, J.G., Smith, A.G., et al., 2004) and The CONCISE GEOLOGIC TIME SCALE (Ogg, J.G., Ogg, G., and Gradstein, F.M., 2008) Paleozoic Sponsored, in part, by: Precambrian ICS Based, in part, on: CENOZOIC-MESOZOIC BIOCHRONOSTRATIGRAPHY: JAN HARDENBOL, JACQUES THIERRY, MARTIN B. FARLEY, THIERRY JACQUIN, PIERRE-CHARLES DE GRACIANSKY, AND PETER R. VAIL,1998. Mesozoic and Cenozoic Sequence Chronostratigraphic Framework of European Basins in: De Graciansky, P.- C., Hardenbol, J., Jacquin, Th., Vail, P. R., and Farley, M. B., eds.; Mesozoic and Cenozoic Sequence Stratigraphy of European Basins, SEPM Special Publication 60. Standard Geo- Ammonites Sequences Ammonites Sequences Smaller Benthic Foraminifers Larger Benthic Foraminifers Calcareous Nannofossils Dinoflagellate Cysts Radiolarians Belemnites Brachiopods Ostracodes Charophytes Stage Age Chronostratigraphy magnetic North Atlantic Tethys Age Polarity Sequences Boreal Sequences T-R Major T-R Period Epoch Stage Substage Boreal Boreal T-R Cycles Tethyan Global, Tethyan Cycles Cycles Zones Tethyan markers Zones Zonal Markers Zonal Markers Zones Boreal Zonal Markers Other Boreal Nannofossils Zones Zonal Markers Other Dinocysts Tethyan Dinocysts Zones Zonal Markers NW Europe Boreal Tethyan Boreal Ostracodes Tethyan Ostracodes Markers Lt. Bajoc. Strenoceras niortense Teloceras banksi Strenoceras niortense Teloceras banksi DSJ14 Lithodinia valensii Belemnopsis Lissajouthyris Lissajouthyris [un-named] 169.8 Stephanolithion speciosum Mancodinium semitabulatum, 169.8 L. galeata mg P., speciosum Durotrigia daveyi, Phallocysta apiciconus matisconensis matisconensis Glyptocythere scitula Teloc. blagdeni Bj3 Telo. blagdeni Bj3 NJ10 Andromeda depressa, G. -

(Middle Jurassic): a Key Interval in the Early Mesozoic Phytoplankton Radiation
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by NERC Open Research Archive 1 The Bajocian (Middle Jurassic): a key interval in the early Mesozoic phytoplankton radiation 2 3 Nickolas J. Wiggan a.b*, James B. Riding b, Robert A. Fensome c, Emanuela Mattioli d 4 5 Insert addresses here 6 7 ABSTRACT 8 Dinoflagellates and coccolithophores are two of the most important groups of phytoplankton 9 in the modern oceans. These groups originated in the Triassic and radiated through the early 10 Mesozoic, rising to ecological prominence. Within this long-term radiation, important short- 11 term intervals of evolutionary and ecological change can be recognised. The Bajocian 12 (Middle Jurassic, ~170–168 Ma) was characterised by an important ecological transition 13 within the coccolithophores, and the radiation of one of the principal families of cyst-forming 14 dinoflagellates, the Gonyaulacaceae. During the Early Bajocian, the coccolith genus 15 Watznaueria diversified and expanded ecologically to dominate coccolith floras, a situation 16 which continued for the remainder of the Mesozoic. This pattern was paralleled within 17 dinoflagellate cyst floras by the ecological dominance of the genus Dissiliodinium in the mid- 18 palaeolatitudes. These phenomena appear to be linked to a positive carbon isotope shift, and 19 an interval of enhanced productivity driven by a shift to a more humid climate, enhanced 20 continental weathering and nutrient flux. The latest Early Bajocian to earliest Bathonian was 21 then characterised by the rapid increase in diversity of dinoflagellate cysts within the family 22 Gonyaulacaceae. Through this interval, the Gonyaulacaceae transitioned from being a 23 relatively minor component of dinoflagellate cyst floras, to becoming one of the prominent 24 groups of cyst-forming dinoflagellates, which has persisted to the Holocene. -

Ammonites from the Latest Aalenian–Earliest Bathonian of La Baume (Castellane Area, SE France): Palaeontology and Biostratigraphy
1661-8726/08/030563-16 Swiss J. Geosci. 101 (2008) 563–578 DOI 10.1007/s00015-008-1297-6 Birkhäuser Verlag, Basel, 2008 Ammonites from the latest Aalenian–earliest Bathonian of La Baume (Castellane area, SE France): palaeontology and biostratigraphy KENNETH DE BAETS 1, 2, *, FABRIZIO CECCA 3, MYETTE GUIOMAR 4 & JACQUES VERNIERS 1 Key words: ammonites, Aalenian–Bathonian interval, France, biodiversity, taxonomy, biostratigraphy ABSTRACT Middle Jurassic strata are naturally exposed around the village called La (Parkinsoni Zone) and probably one lower Bathonian Zone (Zigzag Zone) Baume, near Castellane (Alpes de Haute Provence, SE France). We have re- were found. A major gap of both the Niortense Zone and the Garantiana alized both a detailed log and a bed-by-bed sampling for ammonite biostra- Zone, which was not previously described, was detected at the boundary be- tigraphy in a 68 metres thick succession of subpelagic marls and limestones tween members 2 and 3. The main palaeontological interest of the ammonite (“Calcaires à Zoophycos”) that spans the Middle Jurassic from uppermost fauna from La Baume is the richness and diversity of the family Sonniniidae, Aalenian to lowermost Bathonian. The subpelagic succession can be roughly which is the subject of a systematic study and figurated along with some bio- subdivided into three members. Ammonites from the Upper Aalenian Con- stratigraphically significant forms. Biostratigraphical results and open prob- cavum Zone, all Lower Bajocian Zones (Discites Zone, Laeviuscula Zone lems are discussed. including Ovale Subzone, Humpriesianum Zone), one Upper Bajocian Zone Introduction biostratigraphical subdivisions of the geological formations ex- posed. The Jurassic successions in the area of the Réserve Géologique des Alpes de Haute Provence are widely known because of Geological setting the excellent quality of the exposures, the abundance of am- monoids and the easy access. -

Iterative Evolution of Middle Jurassic Ammonite Faunas
Iterative evolution of Middle Jurassic ammonite faunas ULF EAYER AND GEORGE R. McGHEE. JR Bayer, U. & McGhee, G. R.. Jr. 198401 15: Iterative evolution of Middle Jurassic ammonite Faunas LETHAIA [Fossil-Lagerstatten Nr. 581. Lethaiu. Vol. 17, 1-16. Oslo. ISSN 00?4116J. The phenomenon of iterative evolution of homeomorphs in identical temporal sequences is exhibitcd by Aalenian and Bajocian ammonite faunas in the south German depositional basin. Within each evolution- ary cycle, inflated, evolute, and ornamented shells with complex suture lincs are successively replaced with discoidal, involute, smooth shells with simpler sutures. Three distinct and identical cycles of morphological change occur, and involve ammonites belonging to four different familialisubfamilial groups: the Leioceratinae, Graphoceratinae. Hammatoceratidae, and Sonniniidae. Geometric and statis- tical analyses of morphological change within each of these higher taxa reveal remarkahle iteration in both the pattern and proportion of change with time, The iterative morphological cycles were not driven by orthogenesis or anagenetic transformation within lineages, as has been previously suggested. A clear correlation exists between the successive ammonite faunas and repeated cyclical changes in the physical marine environment of the depositional basin. 0 Ammonoidea, Aulenian, Bajocian, south Germany, iterative er,olution. Ulf Buyer, Imtitut fur Geologie und Pulaontologie der Universitat Tubingen, Sigu,urt.stra/.k 10, 0-7400 Tubingen. F. R. Germany; George R. McGhee, Jr., Institut fur Geologie und Paluontologie der Unirwsitat Tubingen, Sigwurtsirufle 10, 0-7400 Tubingen, F. R. Germany; presenr address: Department of Geological Sciences, Wright Geological Lahorutory. Kutgers Univer.siry, New, Rrunswick. NewJersey 08403 U.S.A.; 4th February, 1983. Iterative evolution of morphologic types - the Evolution of the basin phenomenon of heterochronous homeomorphy - is well known to ammonite workers (Arkell In this section we give the palaeogeographical 1957; Kennedy & Cobban 1977). -

Towards an Integrated Jurassic Biostratigraphy for Eastern Greenland
Volumina Jurassica, 2015, Xiii (1): 43–64 Doi: 10.5604/17313708 .1148657 Towards an integrated Jurassic biostratigraphy for eastern Greenland Simon R.A. KELLY 1, F. John GREGORY 2, William BRAHAM 3, Dominic P. STROGEN 4, Andrew G. WHITHAM 1 Key words: Jurassic, integrated biostratigraphy, ammonites, palynology, micropalaeontology, Greenland. Abstract. The thick and relatively complete Jurassic succession of eastern Greenland provides a unique biostratigraphic record for the North Atlantic region. The main biostratigraphic control for the succession has been provided by molluscs, especially ammonites and to a lesser extent by bivalves and belemnites. The late John Callomon and colleagues recognised 93 Boreal ammonite-bearing horizons in the Mid to Late Jurassic. This provides a reliable backbone to the biostratigraphy of these strata, prompting a palynological colleague to com- ment that they are the “Policemen of Jurassic Stratigraphy”. Other biostratigraphically significant microfossil and palynological groups, can be calibrated against this standard, but on their own cannot achieve the same precision. The Early Jurassic of eastern Greenland does not have such fine control as later parts of the period. No single biostratigraphic group can be used successfully throughout the interval, and there are only three significant ammonite faunas during this period. Reliance on various dif- ferent organisms is necessary to cope with the changing range of marine to non-marine environments. CASP field-work from 1990 to 2012 has resulted in the collection of much biostratigraphic material. In this article, published data are sum- marised together with previously unpublished data in the form of a unified table. The integrated chart shows detailed columns for the whole eastern Greenland Jurassic. -

Jurassic Gastropod Faunas of Central Saudi Arabia
GeoArabia, Vol. 6, No. 1, 2001 Gulf PetroLink, Bahrain Jurassic Gastropod Faunas, Central Saudi Arabia Jurassic Gastropod Faunas of Central Saudi Arabia Jean-Claude Fischer Laboratoire de Paléontologie du Muséum national d’Histoire naturelle, Paris, France Yves-Michel Le Nindre, Jacques Manivit and Denis Vaslet Bureau de Recherches Géologiques et Minières, Orléans, France ABSTRACT Mapping of Phanerozoic rocks at 1:250,000 scale by joint teams from the Saudi Arabian Deputy Ministry for Mineral Resources and the Bureau de Recherches Géologiques et Minières since 1980 has covered most of the Jurassic outcrops in central Saudi Arabia. Stratigraphic, sedimentologic and paleogeographic studies provided a precise framework for collected gastropod faunas that could be calibrated against ammonite zones and sequence-stratigraphic zones. Of more than 600 samples collected, about 440 gastropod specimens could be determined on at least a generic level. Their age range is from Bajocian to Oxfordian–Kimmeridgian. They correspond to 26 genera and 35 species from the Euomphalidae, Ataphridae, Pseudomelaniidae, Coelostylinidae, Procerithiidae, Nerineidae, Purpurinidae, Aporrhaidae, Naticidae, Acteonidae, Retusidae, and Akeridae families. Twelve species are new, and three (Kosmomphalus and Bifidobasis in the Euomphalidae, and Striatoonia in the Pseudomelaniidae) were proposed for new taxa of generic or subgeneric rank. Most of the identified species are of Middle Jurassic age, mainly Bathonian and Callovian and only six are Late Jurassic. All species are typical of an internal platform environment (upper infralittoral), in a lagoonal to back-reef setting, but some also colonized the external platform in the lower infralittoral fore-reef zone. Paleogeographically, most of the species are related to European and Sinai faunas; only seven are equivalent to North African or East African faunas, and one only was reported from Madagascar. -
The Middle Jurassic Palynostratigraphy of the Northern Lusitanian Basin, Portugal
1 The Middle Jurassic palynostratigraphy of the northern Lusitanian Basin, Portugal 2 3 Vânia F. Correia1,2, James B. Riding3, Maria Helena Henriques4, Paulo Fernandes1, 4 Zélia Pereira2, and Nickolas J. Wiggan3,5 5 6 With 10 figures 7 8 Authors’ addresses: 9 1 Corresponding author: CIMA – Centro de Investigação Marinha e Ambiental, Universidade 10 do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal 11 2 LNEG, Rua da Amieira, 4465-965 S. Mamede de Infesta, Portugal 12 3 British Geological Survey, Environmental Science Centre, Keyworth, Nottingham NG12 13 5GG, UK 14 4 Department of Earth Sciences and Centre of Geosciences, Faculty of Science and 15 Technology, University of Coimbra, Rua Sílvio Lima, 3030-790 Coimbra, Portugal 16 5 Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge CB2 17 3EQ, UK 18 * Corresponding author: [email protected] 19 20 Abstract. A composite largely Middle Jurassic succession spanning the Toarcian–Aalenian 21 transition to the lowermost Bathonian exposed at Cabo Mondego and São Gião in the 22 northern Lusitanian Basin, western Portugal, was examined palynologically. The 129 samples 23 are correlated to ammonite biozones (ABs) spanning Pleydellia aalensis to Zigzagiceras 24 zigzag. The Cabo Mondego succession comprises the type section of the Cabo Mondego 25 Formation and spans virtually the entire interval studied. This is a significant interval because 26 it includes the Global Stratotype Section and Point (GSSP) and the Auxiliary Stratigraphical 27 Section and Point (ASSP) for the Bajocian and Bathonian stages respectively. The Cabo 28 Mondego Formation largely yielded relatively abundant palynomorph associations in the 68 1 29 productive samples recovered.