Towards an Integrated Jurassic Biostratigraphy for Eastern Greenland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Post-Carboniferous Stratigraphy, Northeastern Alaska by R
Post-Carboniferous Stratigraphy, Northeastern Alaska By R. L. DETTERMAN, H. N. REISER, W. P. BROSGE,and]. T. DUTRO,JR. GEOLOGICAL SURVEY PROFESSIONAL PAPER 886 Sedirnentary rocks of Permian to Quaternary age are named, described, and correlated with standard stratigraphic sequences UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1975 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress Cataloging in Publication Data Detterman, Robert L. Post-Carboniferous stratigraphy, northeastern Alaska. (Geological Survey Professional Paper 886) Bibliography: p. 45-46. Supt. of Docs. No.: I 19.16:886 1. Geology-Alaska. I. Detterman, Robert L. II. Series: United States. Geological Survey. Professional Paper 886. QE84.N74P67 551.7'6'09798 74-28084 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Stock Number 024-001-02687-2 CONTENTS Page Page Abstract __ _ _ _ _ __ __ _ _ _ _ _ _ _ _ _ _ _ _ __ __ _ _ _ _ _ _ __ __ _ _ __ __ __ _ _ _ _ __ 1 Stratigraphy__:_Continued Introduction __________ ----------____ ----------------____ __ 1 Kingak Shale ---------------------------------------- 18 Purpose and scope ----------------------~------------- 1 Ignek Formation (abandoned) -------------------------- 20 Geographic setting ------------------------------------ 1 Okpikruak Formation (geographically restricted) ________ 21 Previous work and acknowledgments ------------------ 1 Kongakut Formation ---------------------------------- -

The Jurassic Pleurotomarioidean Gastropod Laevitomaria and Its Palaeobiogeographical History
The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history ROBERTO GATTO, STEFANO MONARI, JÁNOS SZABÓ, and MARIA ALESSANDRA CONTI Gatto, R., Monari, S., Szabó, J., and Conti, M.A. 2015. The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history. Acta Palaeontologica Polonica 60 (1): 217–233. The genus Laevitomaria is reviewed and its palaeobiogeographical history is reconstructed based on the re-examination of its type species L. problematica, the study of material stored at the National Natural History Museum of Luxembourg, and an extensive review of the literature. The systematic study allows ascribing to Laevitomaria a number of Jurassic species from the western European region formerly included in other pleurotomariid genera. The following new combi- nations are proposed: Laevitomaria allionta, L. amyntas, L. angulba, L. asurai, L. daityai, L. fasciata, L. gyroplata, L. isarensis, L. joannis, L. repeliniana, L. stoddarti, L. subplatyspira, and L. zonata. The genus, which was once considered as endemic of the central part of the western Tethys, shows an evolutionary and palaeogeographical history consider- ably more complex than previously assumed. It first appeared in the Late Sinemurian in the northern belt of the central western Tethys involved in the Neotethyan rifting, where it experienced a first radiation followed by an abrupt decline of diversity in the Toarcian. Species diversity increased again during Toarcian–Aalenian times in the southernmost part of western European shelf and a major radiation occurred during the Middle Aalenian to Early Bajocian in the northern Paris Basin and southern England. After a latest Bajocian collapse of diversity, Laevitomaria disappeared from both the central part of western Tethys and the European shelf. -
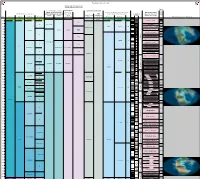
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

Text of Draft
Reconnaissance bedrock geologic map for the northern Alaska Peninsula area, southwest Alaska Including the Dillingham, Iliamna, Lake Clark, Taylor Mountains and the western part of the Kenai and Seldovia 1:250,000-scale quadrangles Compiled by Frederic H. Wilson, Robert B. Blodgett, Charles D. Blomé, Solmaz Mohadjer, Cindi C. Preller, Edward P. Klimasauskas, Bruce M. Gamble, and Warren L. Coonrad DISCLAIMER This report is preliminary and has not been reviewed for conformity with U.S. Geological Survey editorial standards or with the North American Stratigraphic Code. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. This World-Wide-Web publication was prepared by an agency of the United States Government. Neither the United States Government nor any agency thereof, nor any of their employees, makes any warranty, expressed or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed in this report, or represents that its use would not infringe privately owned rights. Reference therein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not necessarily constitute or imply its endorsement, recommendation, or favoring by the United States Government or any agency thereof. Although all data and software published on this Web-site have been used by the USGS, no warranty, expressed or implied, is made by the USGS as to the accuracy of the data and related materials and (or) the functioning of the software. The act of distribution shall not constitute any such warranty, and no responsibility is assumed by the USGS in the use of this data, software, or related materials. -

Malcolm T. Sanders, Jérémie Bardin, Mohammed Benzaggagh & Fabrizio Cecca
Early Toarcian (Jurassic) belemnites from northeastern Gondwana (South Riffian ridges, Morocco) Malcolm T. Sanders, Jérémie Bardin, Mohammed Benzaggagh & Fabrizio Cecca Paläontologische Zeitschrift Scientific Contributions to Palaeontology ISSN 0031-0220 Paläontol Z DOI 10.1007/s12542-013-0214-0 1 23 Your article is protected by copyright and all rights are held exclusively by Springer- Verlag Berlin Heidelberg. This e-offprint is for personal use only and shall not be self- archived in electronic repositories. If you wish to self-archive your article, please use the accepted manuscript version for posting on your own website. You may further deposit the accepted manuscript version in any repository, provided it is only made publicly available 12 months after official publication or later and provided acknowledgement is given to the original source of publication and a link is inserted to the published article on Springer's website. The link must be accompanied by the following text: "The final publication is available at link.springer.com”. 1 23 Author's personal copy Pala¨ontol Z DOI 10.1007/s12542-013-0214-0 RESEARCH PAPER Early Toarcian (Jurassic) belemnites from northeastern Gondwana (South Riffian ridges, Morocco) Malcolm T. Sanders • Je´re´mie Bardin • Mohammed Benzaggagh • Fabrizio Cecca Received: 3 May 2013 / Accepted: 30 October 2013 Ó Springer-Verlag Berlin Heidelberg 2013 Abstract A belemnite fauna collected in the lowermost post-date the earliest Toarcian Polymorphum—Tenuicost- Toarcian succession that crops out near Moulay Idriss atum Chronozone. However, records of Early Jurassic be- (northern Morocco) is studied in this article. This is the first lemnites are still too sparse to recognize the establishment palaeontological study of Early Toarcian belemnites from of provincialism and the timing of its onset. -

The British Toarcian (Lower Jurassic) Belemnites P
THE BRITISH TOARCIAN (LOWER JURASSIC) BELEMNITES P. DOYLE PART 1 Pages 1-49; Plates 1-17 MONOGRAPH OF THE PALAE ON TO GRAPHICAL SOCIETY THE BRITISH TOARCIAN (LOWER JURASSIC) BELEMNITES P. DOYLE PART 1 Pages 1-49: Plates 1-17 © THE PALAE ONTO GRAPHICAL SOCIETY • LONDON December 1990 http://jurassic.ru/ The Palaeontographical Society issues an annual volume of serially numbered publications; these may either be a single complete monograph or part of a continuing monograph. Publication No. 584, issued as part of Volume 144 for 1990 Recommended reference to this publication: DOYLE, P. 1990. The British Toarcian (Lower Jurassic) belemnites. Part 1. Monograph of the Palaeontographical Society London: 1-49, pis 1-17. (Publ. No. 584, part of vol. 144 for 1990). ABSTRACT Part 1 includes sections on the history of previous research, belemnite morphology and the stratigraphy of the British Toarcian outcrop. Five Toarcian belemnite biozones are proposed. 22 species belonging to 4 genera of the Belemnitidae d'Orbigny are described. RESUME La premiere partie comprend l'histoire des recherches, la morphologie des belemnites, et la stratigraphie de Paffleurement Toarcien britannique. Cinq biozones a belemnites sont proposees. 22 especes sont decrites; elles appartiennent a 4 genres de Belemnitidae d'Orbigny. ZUSAMMENFASSUNG Teil 1 enthalt Abschnitte iiber die Geschichte der bisherigen Forschung, die Mor phologie der Belemniten und die Stratigraphie des britischen Toarc-Ausstrichs. Fiinf Belemniten-Biozonen werden im Toarc vorgeschlagen. 22 Arten aus 4 Gattungen der Belemnitidae d'Orbigny werden beschrieben. TOAPCKHE 6EJIEMHHTM BEJMKO6PHTAHHH. PE3K>ME HACTB 1. RIEPBAS NACTB BKJNONAET RJIABW NO HCTOPHH HCAIEFLOBAMRII, MOP<J>OJIORHH 6EJIEMHHT0B H CTPARAR- PA<}>HN 6PHTAHCKHX TOAPCKHX OTJIOACEHHH. -

Geology of the Shepton Mallet Area (Somerset)
Geology of the Shepton Mallet area (Somerset) Integrated Geological Surveys (South) Internal Report IR/03/94 BRITISH GEOLOGICAL SURVEY INTERNAL REPORT IR/03/00 Geology of the Shepton Mallet area (Somerset) C R Bristow and D T Donovan Contributor H C Ivimey-Cook (Jurassic biostratigraphy) The National Grid and other Ordnance Survey data are used with the permission of the Controller of Her Majesty’s Stationery Office. Ordnance Survey licence number GD 272191/1999 Key words Somerset, Jurassic. Subject index Bibliographical reference BRISTOW, C R and DONOVAN, D T. 2003. Geology of the Shepton Mallet area (Somerset). British Geological Survey Internal Report, IR/03/00. 52pp. © NERC 2003 Keyworth, Nottingham British Geological Survey 2003 BRITISH GEOLOGICAL SURVEY The full range of Survey publications is available from the BGS Keyworth, Nottingham NG12 5GG Sales Desks at Nottingham and Edinburgh; see contact details 0115-936 3241 Fax 0115-936 3488 below or shop online at www.thebgs.co.uk e-mail: [email protected] The London Information Office maintains a reference collection www.bgs.ac.uk of BGS publications including maps for consultation. Shop online at: www.thebgs.co.uk The Survey publishes an annual catalogue of its maps and other publications; this catalogue is available from any of the BGS Sales Murchison House, West Mains Road, Edinburgh EH9 3LA Desks. 0131-667 1000 Fax 0131-668 2683 The British Geological Survey carries out the geological survey of e-mail: [email protected] Great Britain and Northern Ireland (the latter as an agency service for the government of Northern Ireland), and of the London Information Office at the Natural History Museum surrounding continental shelf, as well as its basic research (Earth Galleries), Exhibition Road, South Kensington, London projects. -

Palaeoecology and Palaeoenvironments of the Middle Jurassic to Lowermost Cretaceous Agardhfjellet Formation (Bathonian–Ryazanian), Spitsbergen, Svalbard
NORWEGIAN JOURNAL OF GEOLOGY Vol 99 Nr. 1 https://dx.doi.org/10.17850/njg99-1-02 Palaeoecology and palaeoenvironments of the Middle Jurassic to lowermost Cretaceous Agardhfjellet Formation (Bathonian–Ryazanian), Spitsbergen, Svalbard Maayke J. Koevoets1, Øyvind Hammer1 & Crispin T.S. Little2 1Natural History Museum, University of Oslo, P.O. Box 1172 Blindern, 0318 Oslo, Norway. 2School of Earth and Environment, University of Leeds, Leeds LS2 9JT, United Kingdom. E-mail corresponding author (Maayke J. Koevoets): [email protected] We describe the invertebrate assemblages in the Middle Jurassic to lowermost Cretaceous of the Agardhfjellet Formation present in the DH2 rock-core material of Central Spitsbergen (Svalbard). Previous studies of the Agardhfjellet Formation do not accurately reflect the distribution of invertebrates throughout the unit as they were limited to sampling discontinuous intervals at outcrop. The rock-core material shows the benthic bivalve fauna to reflect dysoxic, but not anoxic environments for the Oxfordian–Lower Kimmeridgian interval with sporadic monospecific assemblages of epifaunal bivalves, and more favourable conditions in the Volgian, with major increases in abundance and diversity of Hartwellia sp. assemblages. Overall, the new information from cores shows that abundance, diversity and stratigraphic continuity of the fossil record in the Upper Jurassic of Spitsbergen are considerably higher than indicated in outcrop studies. The inferred life positions and feeding habits of the benthic fauna refine our understanding of the depositional environments of the Agardhfjellet Formation. The pattern of occurrence of the bivalve genera is correlated with published studies of Arctic localities in East Greenland and northern Siberia and shows similarities in palaeoecology with the former but not the latter. -

A Key Interval in the Early Mesozoic Phytoplankton Radiation
1 The Bajocian (Middle Jurassic): a key interval in the early Mesozoic phytoplankton radiation 2 3 Nickolas J. Wiggan a.b*, James B. Riding b, Robert A. Fensome c, Emanuela Mattioli d 4 5 Insert addresses here 6 7 ABSTRACT 8 Dinoflagellates and coccolithophores are two of the most important groups of phytoplankton 9 in the modern oceans. These groups originated in the Triassic and radiated through the early 10 Mesozoic, rising to ecological prominence. Within this long-term radiation, important short- 11 term intervals of evolutionary and ecological change can be recognised. The Bajocian 12 (Middle Jurassic, ~170–168 Ma) was characterised by an important ecological transition 13 within the coccolithophores, and the radiation of one of the principal families of cyst-forming 14 dinoflagellates, the Gonyaulacaceae. During the Early Bajocian, the coccolith genus 15 Watznaueria diversified and expanded ecologically to dominate coccolith floras, a situation 16 which continued for the remainder of the Mesozoic. This pattern was paralleled within 17 dinoflagellate cyst floras by the ecological dominance of the genus Dissiliodinium in the mid- 18 palaeolatitudes. These phenomena appear to be linked to a positive carbon isotope shift, and 19 an interval of enhanced productivity driven by a shift to a more humid climate, enhanced 20 continental weathering and nutrient flux. The latest Early Bajocian to earliest Bathonian was 21 then characterised by the rapid increase in diversity of dinoflagellate cysts within the family 22 Gonyaulacaceae. Through this interval, the Gonyaulacaceae transitioned from being a 23 relatively minor component of dinoflagellate cyst floras, to becoming one of the prominent 24 groups of cyst-forming dinoflagellates, which has persisted to the Holocene. -

The Zoology of East Greenland
/V ^^^tAx^^T^' MEDDELELSER OM GR0NLAND UDGIVNE AF ^ KOMMISSIONEN FOR VIDENSKABELIGE UNDERS0GELSERIGR0NLAND BD. 126 • NR. 6 THE ZOOLOGY OF EAST GREENLAND Edited by M. Degerbel, Ad. S. Jensen, R. Sparck and G. Thorson, Dr. phil. Professor, Dr. phil. Professor, Dr. phil. Dr. phil. in Cooperation with the Editorial Committee of »MeddeleIser om GronIand«. DECAPOD CRUSTACEANS BY P. E. HEEGAARD WITH 27 FIGURES IN THE TEXT 't! % K0BENHAVN C. A. REITZELS FORLAG BIANCO LUNOS BOGTRYKKKRI A/S 1941 Pris: Kr. 3.50. MEDDELELSER OM GR0NLAND UDGIVNE AF KOMMISSIONEN FOR VIDENSKABELIGE UNDERS0GELSER I GR0NLAND BD. 121 • NR. 6 THE ZOOLOGY OF EAST GREENLAND DECAPOD CRUSTACEANS BY P. E. HEEGAARD WITH 27 FIGURES IN THE TEXT K0BENHAVN C. A. REITZELS FORLAG BIANCO LUNOS BOGTRYKKERI A/S 1941 CONTENTS Pa Re Introduction 5 Brachyura Hyas coaretains Anornura Lithode.s- maja — grimaldii Paralomis spectabilis — bouvicri '5 Eupagurus pubescens !*"> Munida lenuimana. Galacanta roslrata Munidopsis eurriroslra 1 — si His Macrura 20 Polycheles nanus Sclerocra.ngon jero.t: 20 — borcas 24 Neetocrangon lar 28 Sabinea, hystri.r sepleincannala 31 Pont o phi I us norvegieus 34 Glyphocrangon sculptus Spirontocaris gainiardu — spin us 39 — lilijeborgii 42 — turgida 42 — polar is 45 groenlandiea 47 Bythocaris payeri 50 — leucopis °2 — simplicirostris 53 Pandalus boreahs 54 — propinquus 5(> Pasiphae tarda. 57 Hymenodora glacial is 58 Amalopeneus elegans 59 Sergestes arclicus "0 General remarks Literature INTRODUCTION The present paper comprises an account of the Crustacean Decapods so far found off the coast of East Greenland. Tt is primarily based on collections made by Danish Expeditions during the last few years, amongst which can be mentioned: ,,Treaarsexpeditionen til Christian d. -

Late Jurassic Ammonites from Alaska
Late Jurassic Ammonites From Alaska GEOLOGICAL SURVEY PROFESSIONAL PAPER 1190 Late Jurassic Ammonites From Alaska By RALPH W. IMLAY GEOLOGICAL SURVEY PROFESSIONAL PAPER 1190 Studies of the Late jurassic ammonites of Alaska enables fairly close age determinations and correlations to be made with Upper Jurassic ammonite and stratigraphic sequences elsewhere in the world UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1981 UNITED STATES DEPARTMENT OF THE INTERIOR JAMES G. WATT, Secretary GEOLOGICAL SURVEY Dallas L. Peck, Director Library of Congress catalog-card No. 81-600164 For sale by the Distribution Branch, U.S. Geological Survey, 604 South Pickett Street, Alexandria, VA 22304 CONTENTS Page Page Abstract ----------------------------------------- 1 Ages and correlations ----------------------------- 19 19 Introduction -------------------------------------- 2 Early to early middle Oxfordian -------------- Biologic analysis _________________________________ _ 14 Late middle Oxfordian to early late Kimmeridgian 20 Latest Kimmeridgian and early Tithonian _____ _ 21 Biostratigraphic summary ------------------------- 14 Late Tithonian ______________________________ _ 21 ~ortheastern Alaska ------------------------- 14 Ammonite faunal setting -------------------------- 22 Wrangell Mountains -------------------------- 15 Geographic distribution ---------------------------- 23 Talkeetna Mountains ------------------------- 17 Systematic descriptions ___________________________ _ 28 Tuxedni Bay-Iniskin Bay area ----------------- 17 References -

Constructional Morphology of the Shell/Ligament System in Opisthogyrate Rostrate Bivalves J
Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 106, 221–227, 2017 Constructional morphology of the shell/ligament system in opisthogyrate rostrate bivalves J. Echevarrı´a, S. E. Damborenea and M. O. Mancen˜ido CONICET – Museo de La Plata, Paseo del Bosque s/n, (1900) La Plata, Buenos Aires province, Argentina. Email: [email protected] ABSTRACT: The bivalve ligament provides the thrust for shell opening, acting as the resistance in a lever system against which adductor muscle effort is applied. Usually, its outer lamellar layer is subjected to tensile stress, while the inner fibrous layer is compressed, with the pivotal axis located between them. However, opisthogyrate rostrate bivalves display a concave dorsal margin, and both the umbo and the postero-dorsal angle of the shell project dorsally to the ligament, which then fails to act as pivotal axis. Three opisthogyrate rostrate genera of unrelated lineages show somewhat dif- ferent solutions to this morpho-functional challenge. In Cuspidaria (Anomalodesmata), the ligament is internal, subjected only to compression and ventral to the pivotal axis, a thickened periostracum develops, forcing the dorsal margins of the valves to act as pivotal axis, and the posterior parts of the shell’s dorsal margins gape dorsally. In Nuculana (Palaeotaxodonta), the inner layer of the ligament is internal, the outer layer is external but reduced, and some species develop a dorsal ridge parallel to the commissural plane, on a level with the rostrum and acting as pivotal axis. In Pterotrigonia (Palaeoheterodonta) and other rostrate trigoniides, the ligament is external opisthodetic, but is allometrically reduced.