Ammonite Zonal Succession of the Middle Jurassic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Or Early Callovian) Ammonites from Alaska and Montana
Jurassic (Bathonian or Early Callovian) Ammonites From Alaska and Montana By RALPH W. IMLAY SHORTER CONTRIBUTIONS TO GENERAL GEOLOGY GEOLOGICAL SURVEY PROFESSIONAL PAPER 374-C Descr$tions and illustrations of ctphalopods of possible late Middle Jurasric (Bathonian) age UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1962 UNITED STATES DEPARTMENT OF THE INTERIOR STEWART L. UDALL, Secretary GEOLOGICAL SURVEY Thomas B. Nolan, Director For sale by the Superintendent of Documents, U.S. Government Printing Office Washington 25, D.C. CONTENTS Page Page C- 1 Age of the faunas-Continued C- 1 Callovian versus Bathonian in Greenland- - - - _ - - _ - - C-2 Callovian versus Bathonian in Alaska and Montana- -- - Stratigraphic summary- __ --______ _ - - - -- - ---.- -- -.- - - C-2 Paleogeographic considerations- - -_-- -- ---- ---- Cook Inlet region, Alaska -______--------.-.--..--c-2 Summation of the evidence- - - _._ _ - _ _ - - - - - - - - - - - - Iniskin Peninsula-_-_______----.--------~.--C-2 Comparisons with other faunas---------___----------- Peninsula north of Chinitna Bay----- __._ _ _._ - C-3 \Vestern interior of Canada- - - -- -- -____------- --- Talkeetna Mountains ----___-_ - - -- ---- - - -- - -- C-3 Arctic region-_-_---___-_----------------------- Western Montana- - -----__-----------------.---C-5 other regions--__-__-____----------------------- Rocky Mountain front north of the Sun River- (2-5 Geographic distribution ___-___ --- - ---------- ------ -- - Drummond area--- ---_____ _--- -- -.-- ---- -- - C-10 Summary of results- --_-____-_----_---_-_----------- Age ofthe faunas-----------_----------------------- GI0 Systematic descriptions--_ _ _ - _ - - - - - - - - - - - - - - - - - - - - - - - Evidence from Alaska---____________--------------C-10 Literature cited _-_-_---______----------------------- Evidence from Montana --_-_____ --- - - -- .--- --- - - C-12 Index---__--___-_-_------------------------------- ILLUSTRATIONS [Plates 1-3 follow index] PLATE 1. Holcophylloceras, Oecotraustes (Paroecotraustes) ?, and Arctocephalites (Cranocephalites). 2. -

The Middle Jurassic of Western and Northern Europe: Its Subdivisions, Geochronology and Correlations
The Middle Jurassic of western and northern Europe: its subdivisions, geochronology and correlations John H. Callomon The palaeogeographic settings of Denmark and East Greenland during the Middle Jurassic are outlined. They lay in the widespread epicontinental seas that covered much of Europe in the post-Triassic transgression. It was a period of continuing eustatic sea-level rise, with only distant connections to world oceans: to the Pacific, via the narrow Viking Straits between Greenland and Norway and hence the arctic Boreal Sea to the north; and to the subtropical Tethys, via some 1200 km of shelf-seas to the south. The sedimentary history of the region was strongly influenced by two factors: tectonism and climate. Two modes of tectonic movement governed basinal evolution: crustal extension lead- ing to subsidence through rifting, such as in the Viking and Central Grabens of the North Sea; and subcrustal thermal upwelling, leading to domal uplift and the partition of marine basins through emergent physical barriers, as exemplified by the Central North Sea Dome with its associated volcanics. The climatic gradient across the 30º of temperate latitude spanned by the European seas governed biotic diversity and biogeography, finding expression in rock-forming biogenic carbonates that dominate sediments in the south and give way to largely siliciclastic sediments in the north. Geochronology of unrivalled finesse is provided by standard chronostratigraphy based on the biostratigraphy of ammonites. The Middle Jurassic saw the onset of considerable bioprovincial endemisms in these guide-fossils, making it necessary to construct parallel standard zonations for Boreal, Subboreal or NW European and Submediterranean Provinces, of which the NW European zonation provides the primary international standard. -

The Jurassic Pleurotomarioidean Gastropod Laevitomaria and Its Palaeobiogeographical History
The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history ROBERTO GATTO, STEFANO MONARI, JÁNOS SZABÓ, and MARIA ALESSANDRA CONTI Gatto, R., Monari, S., Szabó, J., and Conti, M.A. 2015. The Jurassic pleurotomarioidean gastropod Laevitomaria and its palaeobiogeographical history. Acta Palaeontologica Polonica 60 (1): 217–233. The genus Laevitomaria is reviewed and its palaeobiogeographical history is reconstructed based on the re-examination of its type species L. problematica, the study of material stored at the National Natural History Museum of Luxembourg, and an extensive review of the literature. The systematic study allows ascribing to Laevitomaria a number of Jurassic species from the western European region formerly included in other pleurotomariid genera. The following new combi- nations are proposed: Laevitomaria allionta, L. amyntas, L. angulba, L. asurai, L. daityai, L. fasciata, L. gyroplata, L. isarensis, L. joannis, L. repeliniana, L. stoddarti, L. subplatyspira, and L. zonata. The genus, which was once considered as endemic of the central part of the western Tethys, shows an evolutionary and palaeogeographical history consider- ably more complex than previously assumed. It first appeared in the Late Sinemurian in the northern belt of the central western Tethys involved in the Neotethyan rifting, where it experienced a first radiation followed by an abrupt decline of diversity in the Toarcian. Species diversity increased again during Toarcian–Aalenian times in the southernmost part of western European shelf and a major radiation occurred during the Middle Aalenian to Early Bajocian in the northern Paris Basin and southern England. After a latest Bajocian collapse of diversity, Laevitomaria disappeared from both the central part of western Tethys and the European shelf. -

Non-Invasive Imaging Methods Applied to Neo- and Paleo-Ontological Cephalopod Research
Biogeosciences, 11, 2721–2739, 2014 www.biogeosciences.net/11/2721/2014/ doi:10.5194/bg-11-2721-2014 © Author(s) 2014. CC Attribution 3.0 License. Non-invasive imaging methods applied to neo- and paleo-ontological cephalopod research R. Hoffmann1, J. A. Schultz2, R. Schellhorn2, E. Rybacki3, H. Keupp4, S. R. Gerden1, R. Lemanis1, and S. Zachow5 1Institut für Geologie, Mineralogie und Geophysik, Ruhr Universität Bochum, Universitätsstrasse 150, 44801 Bochum, Germany 2Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Nussallee 8, 53115 Bonn, Germany 3Helmholtz-Zentrum Potsdam, Deutsches GeoForschungsZentrum GFZ Sektion 3.2, Geomechanik und Rheologie, Telegrafenberg, D 429, 14473 Potsdam, Germany 4Institut für Geologische Wissenschaften, Fachrichtung Paläontologie, Freie Universität Berlin, Malteserstrasse 74–100, 12249 Berlin, Germany 5Zuse Institut Berlin, Takustrasse 7, 14195 Berlin, Germany Correspondence to: R. Hoffmann ([email protected]) Received: 28 October 2013 – Published in Biogeosciences Discuss.: 29 November 2013 Revised: 14 March 2014 – Accepted: 29 March 2014 – Published: 22 May 2014 Abstract. Several non-invasive methods are common prac- tially preserved within the surrounding rocks, requires imag- tice in natural sciences today. Here we present how they ing methods that are primarily used in non-destructive test- can be applied and contribute to current topics in cephalo- ing. The conservation of the specimen is of main importance pod (paleo-) biology. Different methods will be compared in using these methods since former techniques used destruc- terms of time necessary to acquire the data, amount of data, tive methods leading to the loss of the specimen or parts accuracy/resolution, minimum/maximum size of objects that of the specimen. -

Correlations of the Jurassic Sediments: Infra-Getic Unit
GEOLO[KI ANALI BALKANSKOGA POLUOSTRVA 67 19–33 BEOGRAD, decembar 2006 ANNALES GÉOLOGIQUES DE LA PÉNINSULE BALKANIQUE BELGRADE, December 2006 Tran-sborder (south-east Serbia/west Bulgaria) correlations of the Jurassic sediments: Infra-Getic Unit 1 2 PLATON TCHOUMATCHENCO , DRAGOMAN RABRENOVI] , 3 4 BARBARA RADULOVI] & VLADAN RADULOVI] Abstract. The Infra-Getic Unit is a palaeogeographic unit, predestined by palaeotectonics. From the point of view of geological heritage, it represents a geosites framework. For the purpose of the correlation, the Serbian sections of Lukanja, Bogorodica Monastery, Rosoma~ and Senokos, as well as the Bulgarian sections of Komshtitsa, Gintsi, and Stanyantsi were used. The Jurassic sediments of the Infra-Getic Unit crop out on the southern slops of the Stara Planina Mountain in east Serbia and west Bulgaria. The Lower Jurassic started with continental and continental-marine sediments (clays and sandstones) (Lukanja clastics and Lukanja coal beds in Serbia and the Tuden Formation in Bulgaria) and continue with Lukanja quartz sandstones (Serbia) and the Kostina Formation (Bulgaria). These sediments are covered by Lukanja brachiopod beds and Lukanja limestones (Serbia) and the Romanov Dol, Ravna and Dolni Loukovit Members of the Ozirovo Formation (Bulgaria) pre- dominantly consist of bioclastic limestones. The sedimentations follow with Lukanja belemnites-gryphaea beds (marls and clayey limestones), which in Bulgaria correspond to the Bukorovtsi Member (also marls and clayey limestones) of the Ozirovo Formation. The Middle Jurassic sedimentation started with black shales with Bossitra alpine. These sediments are individualized in Serbia as Senokos aleurolites and clays and in Bulgaria they are known as the Etropole Formation. In Serbia the section continues with sandstones called Vodeni~ki sandstones of Bajocian age, known in Bulgaria as the Dobrogled Member of the Polaten Formation. -

Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda)
GFF ISSN: 1103-5897 (Print) 2000-0863 (Online) Journal homepage: http://www.tandfonline.com/loi/sgff20 Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda) Harry Mutvei To cite this article: Harry Mutvei (2016): Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda), GFF, DOI: 10.1080/11035897.2016.1227364 To link to this article: http://dx.doi.org/10.1080/11035897.2016.1227364 Published online: 21 Sep 2016. Submit your article to this journal View related articles View Crossmark data Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=sgff20 Download by: [Dr Harry Mutvei] Date: 21 September 2016, At: 11:07 GFF, 2016 http://dx.doi.org/10.1080/11035897.2016.1227364 Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda) Harry Mutvei Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-10405 Stockholm, Sweden ABSTRACT ARTICLE HISTORY The shell wall in Spirula is composed of prismatic layers, whereas the septa consist of lamello-fibrillar nacre. Received 13 May 2016 The septal neck is holochoanitic and consists of two calcareous layers: the outer lamello-fibrillar nacreous Accepted 23 June 2016 layer that continues from the septum, and the inner pillar layer that covers the inner surface of the septal KEYWORDS neck. The pillar layer probably is a structurally modified simple prisma layer that covers the inner surface of Siphuncular structures; the septal neck in Nautilus. The pillars have a complicated crystalline structure and contain high amount of connecting rings; Spirula; chitinous substance. -

Organic Carbon Isotope Chemostratigraphy of Late Jurassic Early Cretaceous Arctic Canada
University of Plymouth PEARL https://pearl.plymouth.ac.uk Faculty of Science and Engineering School of Geography, Earth and Environmental Sciences Finding the VOICE: organic carbon isotope chemostratigraphy of Late Jurassic Early Cretaceous Arctic Canada Galloway, JM http://hdl.handle.net/10026.1/15324 10.1017/s0016756819001316 Geological Magazine Cambridge University Press (CUP) All content in PEARL is protected by copyright law. Author manuscripts are made available in accordance with publisher policies. Please cite only the published version using the details provided on the item record or document. In the absence of an open licence (e.g. Creative Commons), permissions for further reuse of content should be sought from the publisher or author. Proof Delivery Form Geological Magazine Date of delivery: Journal and vol/article ref: geo 1900131 Number of pages (not including this page): 15 This proof is sent to you on behalf of Cambridge University Press. Please check the proofs carefully. Make any corrections necessary on a hardcopy and answer queries on each page of the proofs Please return the marked proof within 2 days of receipt to: [email protected] Authors are strongly advised to read these proofs thoroughly because any errors missed may appear in the final published paper. This will be your ONLY chance to correct your proof. Once published, either online or in print, no further changes can be made. To avoid delay from overseas, please send the proof by airmail or courier. If you have no corrections to make, please email [email protected] to save having to return your paper proof. If corrections are light, you can also send them by email, quoting both page and line number. -

Stratigraphic Implications of a New Lower Cretaceous Ammonoid Fauna from the Puez Area (Valanginian – Aptian, Dolomites, Southern Alps, Italy)
Geo.Alp, Vol. 3, S. 55–83, 2006 STRATIGRAPHIC IMPLICATIONS OF A NEW LOWER CRETACEOUS AMMONOID FAUNA FROM THE PUEZ AREA (VALANGINIAN – APTIAN, DOLOMITES, SOUTHERN ALPS, ITALY) Alexander Lukeneder1 & Christian Aspmair2 With 6 figures and 8 plates 1 Natural History Museum, Geological-Palaeontological Department, Burgring 7, A-1010 Wien, Austria, e-mail: [email protected] 2 Prissian 102, I – 39010 Tisens (BZ), Italy Abstract Lower Cretaceous ammonoids (n = 424) were collected at the Puez locality in the Dolomites of Southern Tyrol. The cephalopod fauna from the marly limestones to marls here indicates Late Valanginian to Early Aptian age. The deposition of the marly limestones and marls of this interval occurred during depositional- ly unstable conditions. The underlying Biancone Formation (Maiolica Formation) is of Early Valanginian, whereas the lowermost Rosso Ammonitico is of Jurassic to Berriasian age. The ammonoid fauna consists of 27 different genera, each represented by 1-2 species. The assemblage at the Puez section is dominated by the Phylloceratina (30%) and the Ammonitina (34%). Phyllopachyceras (17%) and Phylloceras (13%) (both Phylloceratina) are the most frequent components, followed by Lytoceras (12%) (Lytoceratina), and Barremites (10%) and Melchiorites (8%) (both Ammonitina). The cephalopod fauna is purely of Mediterranean origin. Zusammenfassung Unterkreide Ammonoideen (424 Exemplare) der Puez Lokalität in den Dolomiten Süd-Tirols wurden unter- sucht. Die Fauna der mergeligen Kalke und Mergel von Puez zeigen ein Alter von Ober-Valanginium bis Unter-Aptium an. Die mergeligen Kalke und Mergel dieses Abschnitts lagerten sich unter instabiler Bedingungen ab. Die unterlagernde Biancone Formation (Maiolica Formation) zeigt Unter-Valanginium an, wogegen die tiefste Formation des Rosso Ammonitico auf Ober-Jura bis Berriasium hindeutet. -

Abelisaurus Comahuensis 321 Acanthodiscus Sp. 60, 64
Index Page numbers in italic denote figure. Page numbers in bold denote tables. Abelisaurus comahuensis 321 structure 45-50 Acanthodiscus sp. 60, 64 Andean Fold and Thrust Belt 37-53 Acantholissonia gerthi 61 tectonic evolution 50-53 aeolian facies tectonic framework 39 Huitrin Formation 145, 151-152, 157 Andes, Neuqu6n 2, 3, 5, 6 Troncoso Member 163-164, 167, 168 morphostructural units 38 aeolian systems, flooded 168, 169, 170, 172, stratigraphy 40 174-182 tectonic evolution, 15-32, 37-39, 51 Aeolosaurus 318 interaction with Neuqu6n Basin 29-30 Aetostreon 200, 305 Andes, topography 37 Afropollis 76 Andesaurus delgadoi 318, 320 Agrio Fold and Thrust Belt 3, 16, 18, 29, 30 andesite 21, 23, 26, 42, 44 development 41 anoxia see dysoxia-anoxia stratigraphy 39-40, 40, 42 Aphrodina 199 structure 39, 42-44, 47 Aphrodina quintucoensis 302 uplift Late Cretaceous 43-44 Aptea notialis 75 Agrio Formation Araucariacites australis 74, 75, 76 ammonite biostratigraphy 58, 61, 63, 65, 66, Araucarioxylon 95,273-276 67 arc morphostructural units 38 bedding cycles 232, 234-247 Arenicolites 193, 196 calcareous nannofossil biostratigraphy 68, 71, Argentiniceras noduliferum 62 72 biozone 58, 61 highstand systems tract 154 Asteriacites 90, 91,270 lithofacies 295,296, 297, 298-302 Asterosoma 86 92 marine facies 142-143, 144, 153 Auca Mahuida volcano 25, 30 organic facies 251-263 Aucasaurus garridoi 321 palaeoecology 310, 311,312 Auquilco evaporites 42 palaeoenvironment 309- 310, 311, Avil6 Member 141,253, 298 312-313 ammonites 66 palynomorph biostratigraphy 74, -
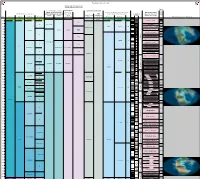
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

Guidebook for the Post-Congress Fieldtrip in the Vocontian Basin, SE France (September 11-13, 2008)
12th Meeting of the International Nannoplankton Association (Lyon, September 7-10, 2008) Guidebook for the post-congress fieldtrip in the Vocontian Basin, SE France (September 11-13, 2008) Emanuela MATTIOLI (special editor), Silvia GARDIN, Fabienne GIRAUD, Davide OLIVERO, Bernard PITTET & Stéphane REBOULET ISBN 978-2-916733-02-9 Dépôt légal à parution Manuscrit en ligne depuis le 7 Septembre 2008 Manuscript online since September 7, 2008 Carnets de Géologie / Notebooks on Geology - Book 2008/01 (CG2008_BOOK_01) Guidebook for the post-congress fieldtrip in the Vocontian Basin, SE France (September 11-13, 2008) Emanuela MATTIOLI (special editor) 1,2, Silvia GARDIN 3, Fabienne GIRAUD 1, Davide OLIVERO 1, Bernard PITTET 1 & Stéphane REBOULET 1 1 UMR 5125 PEPS (CNRS), Université Lyon 1, Campus de la DOUA, Bâtiment Géode, 69622 Villeurbanne Cedex (France) 2 [email protected] 3 UMR 5143 (CNRS), Université Pierre et Marie Curie, Paris 6, 75013 Paris (France) Manuscript online since September 7, 2008 1 Carnets de Géologie / Notebooks on Geology - Book 2008/01 (CG2008_BOOK_01) Chapter 1. The Aalenian-Bajocian northward, with many hiatuses (JAUTÉE, 1984). (Middle Jurassic) of the Digne Basinal facies, represented by marl-limestone alternations, are recorded in the southernmost area areas but in the sector south of the Verdon Davide OLIVERO with the contribution of River a shallow-water platform developed, Emanuela MATTIOLI documented by the occurrence of bioclastic limestones with corals. This development is Geographical and geological context evidence of a progressive extension to the Provence platform southward. The Digne area is located in southeastern France (Fig. 1.1), in the department of "Alpes In the Middle Jurassic, the central part of the de Haute-Provence". -

Schmitz, M. D. 2000. Appendix 2: Radioisotopic Ages Used In
Appendix 2 Radioisotopic ages used in GTS2020 M.D. SCHMITZ 1285 1286 Appendix 2 GTS GTS Sample Locality Lat-Long Lithostratigraphy Age 6 2s 6 2s Age Type 2020 2012 (Ma) analytical total ID ID Period Epoch Age Quaternary À not compiled Neogene À not compiled Pliocene Miocene Paleogene Oligocene Chattian Pg36 biotite-rich layer; PAC- Pieve d’Accinelli section, 43 35040.41vN, Scaglia Cinerea Fm, 42.3 m above base of 26.57 0.02 0.04 206Pb/238U B2 northeastern Apennines, Italy 12 29034.16vE section Rupelian Pg35 Pg20 biotite-rich layer; MCA- Monte Cagnero section (Chattian 43 38047.81vN, Scaglia Cinerea Fm, 145.8 m above base 31.41 0.03 0.04 206Pb/238U 145.8, equivalent to GSSP), northeastern Apennines, Italy 12 28003.83vE of section MCA/84-3 Pg34 biotite-rich layer; MCA- Monte Cagnero section (Chattian 43 38047.81vN, Scaglia Cinerea Fm, 142.8 m above base 31.72 0.02 0.04 206Pb/238U 142.8 GSSP), northeastern Apennines, Italy 12 28003.83vE of section Eocene Priabonian Pg33 Pg19 biotite-rich layer; MASS- Massignano (Oligocene GSSP), near 43.5328 N, Scaglia Cinerea Fm, 14.7 m above base of 34.50 0.04 0.05 206Pb/238U 14.7, equivalent to Ancona, northeastern Apennines, 13.6011 E section MAS/86-14.7 Italy Pg32 biotite-rich layer; MASS- Massignano (Oligocene GSSP), near 43.5328 N, Scaglia Cinerea Fm, 12.9 m above base of 34.68 0.04 0.06 206Pb/238U 12.9 Ancona, northeastern Apennines, 13.6011 E section Italy Pg31 Pg18 biotite-rich layer; MASS- Massignano (Oligocene GSSP), near 43.5328 N, Scaglia Cinerea Fm, 12.7 m above base of 34.72 0.02 0.04 206Pb/238U