Apicoplast Isoprenoid Precursor Synthesis and Molecular Basis Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
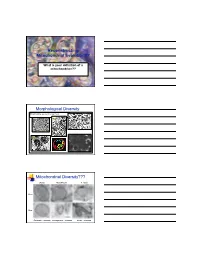
Reconstructing Mitochondrial Evolution?? Morphological Diversity Mitochondrial Diversity???
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial Diversity??? Chicken Neocallimastix T. foetus 100 nm 50 nm Entamoeba - mitosome microsporidian - mitosome Giardia - mitosome Reconstructing Evolution Mitochondrial evolution well established endosymbiotic theory α-proteobacterium - Rickettsia prowazekii Hydrogenosomal evolution No DNA NOW 2 examples Nyctotherus and Blastocystis (MLO) Several proteins similar to mitochondria Mitosome evolution No DNA Few proteins identified similar to mitochondria Origins via Endosymbiosis Current dogma - mitochondria and related Aerobic α-proteobacterium prokaryote gave rise to present day mitochondria. organelles arose just once in evolution Are hydrogenosomes and mitosomes of anaerobic protists derived from the same proto-mitochondrion? Evidence for: accumulating evidence for several proteins that are currently found in mitochondria - Proteins of Fe-S cluster formation. Scenario A Common ancestor facultative Scenario B organism? degenerate mitochondrion invoke lateral gene transfer from anaerobic prokaryotes Hypotheses for Mito Acquisition Mitochondrion-related Organelles 2 way to interpret this summary - can you think of both? Groups with mitochondrial homologues Hjort et al Phil. Trans. R. Soc. B 2010 365, 713-727 Origin of Hydrogenosomes (Mitosomes?) 1. a)Conversion of Mitochondria HYD b)Common ancestor with mitochondria ? HYD 2. Independent Origin α-proteobacterium ? - anaerobic bacterium HYD Organelles - origins and biogenesis Approaches: (1) Conduct phylogenetic analyses of similar proteins Hsp70 Fd Hsp60 Isc subunits (2) Examine protein targeting to the organelle matrix protein targeting membrane protein targeting (3) Characterize membrane/translocation components These components could have evolved as the endosymbiont was converted to organelle. Reveals evolutionary history. -

The Apicoplast: a Review of the Derived Plastid of Apicomplexan Parasites
Curr. Issues Mol. Biol. 7: 57-80. Online journalThe Apicoplastat www.cimb.org 57 The Apicoplast: A Review of the Derived Plastid of Apicomplexan Parasites Ross F. Waller1 and Geoffrey I. McFadden2,* way to apicoplast discovery with studies of extra- chromosomal DNAs recovered from isopycnic density 1Botany, University of British Columbia, 3529-6270 gradient fractionation of total Plasmodium DNA. This University Boulevard, Vancouver, BC, V6T 1Z4, Canada group recovered two DNA forms; one a 6kb tandemly 2Plant Cell Biology Research Centre, Botany, University repeated element that was later identifed as the of Melbourne, 3010, Australia mitochondrial genome, and a second, 35kb circle that was supposed to represent the DNA circles previously observed by microscopists (Wilson et al., 1996b; Wilson Abstract and Williamson, 1997). This molecule was also thought The apicoplast is a plastid organelle, homologous to to be mitochondrial DNA, and early sequence data of chloroplasts of plants, that is found in apicomplexan eubacterial-like rRNA genes supported this organellar parasites such as the causative agents of Malaria conclusion. However, as the sequencing effort continued Plasmodium spp. It occurs throughout the Apicomplexa a new conclusion, that was originally embraced with and is an ancient feature of this group acquired by the some awkwardness (“Have malaria parasites three process of endosymbiosis. Like plant chloroplasts, genomes?”, Wilson et al., 1991), began to emerge. apicoplasts are semi-autonomous with their own genome Gradually, evermore convincing character traits of a and expression machinery. In addition, apicoplasts import plastid genome were uncovered, and strong parallels numerous proteins encoded by nuclear genes. These with plastid genomes from non-photosynthetic plants nuclear genes largely derive from the endosymbiont (Epifagus virginiana) and algae (Astasia longa) became through a process of intracellular gene relocation. -

Reevaluation of the Toxoplasma Gondii and Neospora Caninum Genomes Reveals Misassembly, Karyotype Differences and Chromosomal Rearrangements
bioRxiv preprint doi: https://doi.org/10.1101/2020.05.22.111195; this version posted May 25, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Reevaluation of the Toxoplasma gondii and Neospora caninum genomes reveals misassembly, karyotype differences and chromosomal rearrangements Luisa Berna1, Pablo Marquez1, Andrés Cabrera1, Gonzalo Greif1, María E. Francia2,3,*, Carlos Robello1,4,* AUTHORS AFFILIATIONS 1 Laboratory of Host Pathogen Interactions-Molecular Biology Unit. Institut Pasteur de Montevideo. Montevideo, Uruguay 2 Laboratory of Apicomplexan Biology. Institut Pasteur de Montevideo. Montevideo, Uruguay. 3 Departamento de Parasitología y Micología. Facultad de Medicina-Universidad de la República. Montevideo, Uruguay. 4 Departamento de Bioquímica. Facultad de Medicina-Universidad de la República. Montevideo, Uruguay. *Corresponding authors ABSTRACT Neospora caninum primarily infects cattle causing abortions with an estimated impact of a billion dollars on worldwide economy, annually. However, the study of its biology has been unheeded by the established paradigm that it is virtually identical to its close relative, the widely studied human pathogen, Toxoplasma gondii. By revisiting the genome sequence, assembly and annotation using third generation sequencing technologies, here we show that the N. caninum genome was originally incorrectly assembled under the presumption of synteny with T. gondii. We show that major chromosomal rearrangements have occurred between these species. Importantly, we show that chromosomes originally annotated as ChrVIIb and VIII are indeed fused, reducing the karyotype of both N. -

The NTP Generating Activity of Pyruvate Kinase II Is Critical
RESEARCH ARTICLE The NTP generating activity of pyruvate kinase II is critical for apicoplast maintenance in Plasmodium falciparum Russell P Swift, Krithika Rajaram, Cyrianne Keutcha, Hans B Liu, Bobby Kwan, Amanda Dziedzic, Anne E Jedlicka, Sean T Prigge* Department of Molecular Microbiology and Immunology, Johns Hopkins Bloomberg School of Public Health, Baltimore, United States Abstract The apicoplast of Plasmodium falciparum parasites is believed to rely on the import of three-carbon phosphate compounds for use in organelle anabolic pathways, in addition to the generation of energy and reducing power within the organelle. We generated a series of genetic deletions in an apicoplast metabolic bypass line to determine which genes involved in apicoplast carbon metabolism are required for blood-stage parasite survival and organelle maintenance. We found that pyruvate kinase II (PyrKII) is essential for organelle maintenance, but that production of pyruvate by PyrKII is not responsible for this phenomenon. Enzymatic characterization of PyrKII revealed activity against all NDPs and dNDPs tested, suggesting that it may be capable of generating a broad range of nucleotide triphosphates. Conditional mislocalization of PyrKII resulted in decreased transcript levels within the apicoplast that preceded organelle disruption, suggesting that PyrKII is required for organelle maintenance due to its role in nucleotide triphosphate generation. Introduction *For correspondence: With increasing resistance to current front-line antimalarials, there is a crucial need to find new thera- [email protected] peutic interventions with novel mechanisms of action (Dondorp et al., 2009; Trape, 2001). The api- coplast organelle within the parasite has often been considered as a source of new drug targets Competing interests: The since it is required for blood-stage survival, in addition to possessing evolutionarily distinct biochem- authors declare that no ical pathways that are not present in the human host (Goodman and McFadden, 2013; competing interests exist. -

The Protozoan Nucleus. Molecular and Biochemical Parasitology, 209(1-2), Pp
McCulloch, R., and Navarro, M. (2016) The protozoan nucleus. Molecular and Biochemical Parasitology, 209(1-2), pp. 76-87. (doi:10.1016/j.molbiopara.2016.05.002) This is the author’s final accepted version. There may be differences between this version and the published version. You are advised to consult the publisher’s version if you wish to cite from it. http://eprints.gla.ac.uk/135296/ Deposited on: 22 February 2017 Enlighten – Research publications by members of the University of Glasgow http://eprints.gla.ac.uk *Manuscript Click here to view linked References The protozoan nucleus Richard McCulloch1 and Miguel Navarro2 1. The Wellcome Trust Centre for Molecular Parasitology, Institute of Infection, Immunity and Inflammation, University of Glasgow, Sir Graeme Davis Building, 120 University Place, Glasgow, G12 8TA, U.K. Telephone: 01413305946 Fax: 01413305422 Email: [email protected] 2. Instituto de Parasitología y Biomedicina López-Neyra, Consejo Superior de Investigaciones Científicas (CSIC), Avda. del Conocimiento s/n, 18100 Granada, Spain. Email: [email protected] Correspondence can be sent to either of above authors Keywords: nucleus; mitosis; nuclear envelope; chromosome; DNA replication; gene expression; nucleolus; expression site body 1 Abstract The nucleus is arguably the defining characteristic of eukaryotes, distinguishing their cell organisation from both bacteria and archaea. Though the evolutionary history of the nucleus remains the subject of debate, its emergence differs from several other eukaryotic organelles in that it appears not to have evolved through symbiosis, but by cell membrane elaboration from an archaeal ancestor. Evolution of the nucleus has been accompanied by elaboration of nuclear structures that are intimately linked with most aspects of nuclear genome function, including chromosome organisation, DNA maintenance, replication and segregation, and gene expression controls. -

Mitosomes in Entamoeba Histolytica Contain a Sulfate Activation Pathway
Mitosomes in Entamoeba histolytica contain a sulfate activation pathway Fumika Mi-ichi, Mohammad Abu Yousuf, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki1 Department of Parasitology, National Institute of Infectious Diseases, Tokyo 162-8640, Japan Edited by Andrew Roger, Dalhousie University, Halifax, NS, Canada, and accepted by the Editorial Board October 19, 2009 (received for review June 25, 2009) Hydrogenosomes and mitosomes are mitochondrion-related or- and M. balamuthi lack the ISC system, and instead possess the ganelles in anaerobic/microaerophilic eukaryotes with highly re- nitrogen fixation (NIF) system, which is most likely derived from an duced and divergent functions. The full diversity of their content ancestral nitrogen fixing -proteobacterium by lateral gene transfer and function, however, has not been fully determined. To under- (22, 24). Only 5 proteins have been demonstrated in E. histolytica stand the central role of mitosomes in Entamoeba histolytica,a mitosomes: Cpn60 (8–10, 12), Cpn10 (13), mitochondrial Hsp70 parasitic protozoon that causes amoebic dysentery and liver ab- (11, 15), pyridine nucleotide transhydrogenase (PNT) (2, 8), and scesses, we examined the proteomic profile of purified mitosomes. mitochondria carrier family (MCF, ADP/ATP transporter) (14), Using 2 discontinuous Percoll gradient centrifugation and MS and the central role of mitosomes in E. histolytica remains unknown. analysis, we identified 95 putative mitosomal proteins. Immuno- Analysis of the genome of E. histolytica has not revealed any fluorescence assay showed that 3 proteins involved in sulfate additional information regarding the function of mitosomes and activation, ATP sulfurylase, APS kinase, and inorganic pyrophos- thus, a proteomic analysis of mitosomes seems to be the best phatase, as well as sodium/sulfate symporter, involved in sulfate approach to understand its structure and function (1, 2). -

A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba Histolytica
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Current Biology, Vol. 15, 737–742, April 26, 2005, ©2005 Elsevier Ltd All rights reserved. DOI 10.1016/j.cub.2005.02.068 A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba histolytica Ka Wai Chan,1,5 Dirk-Jan Slotboom,1,5 Sian Cox,3 ses confirm that the Entamoeba ADP/ATP carrier is T. Martin Embley,2,* Olivier Fabre,1 distinct from archetypal mitochondrial ADP/ATP carri- Mark van der Giezen,3,6 Marilyn Harding,1 ers, an observation that is supported by its different David S. Horner,4 Edmund R.S. Kunji,1,* substrate and inhibitor specificity. Because many Gloria León-Avila,3,7 and Jorge Tovar3 functions of yeast and human mitochondria rely on 1Dunn Human Nutrition Unit solutes transported by specialized members of this Medical Research Council family, the Entamoeba mitosome must contain only Hills Road a small subset of these processes requiring adenine Cambridge CB2 2XY nucleotide exchange. United Kingdom 2 School of Biology The Devonshire Building Results and Discussion University of Newcastle upon Tyne Newcastle NE1 7RU Identification of an MCF Homolog United Kingdom on the Entamoeba histolytica Genome 3 School of Biological Sciences We identified a homolog of the mitochondrial carrier Royal Holloway family (MCF) on the Entamoeba histolytica genome (TIGR) University of London with BLAST searches. The translated protein contains a Egham, Surrey TW20 0EX tripartite structure [8], signature motifs (IPR001993), and United Kingdom other features typical of MCF members (Figure 1A). -

Protein Targeting in Parasites with Cryptic Mitochondria
International Journal for Parasitology 37 (2007) 265–272 www.elsevier.com/locate/ijpara Invited review Protein targeting in parasites with cryptic mitochondria Lena Burri, Patrick J. Keeling * Canadian Institute for Advanced Research, Department of Botany, University of British Columbia, 3529-6270 University Boulevard, Vancouver, BC, Canada V6T 1Z4 Received 4 October 2006; received in revised form 5 December 2006; accepted 11 December 2006 Abstract Many highly specialised parasites have adapted to their environments by simplifying different aspects of their morphology or bio- chemistry. One interesting case is the mitochondrion, which has been subject to strong reductive evolution in parallel in several different parasitic groups. In extreme cases, mitochondria have degenerated so much in physical size and functional complexity that they were not immediately recognised as mitochondria, and are now referred to as ‘cryptic’. Cryptic mitochondrion-derived organelles can be classified as either hydrogenosomes or mitosomes. In nearly all cases they lack a genome and all organellar proteins are nucleus-encoded and expressed in the cytosol. The same is true for the majority of proteins in canonical mitochondria, where the proteins are directed to the organelle by specific targeting sequences (transit peptides) that are recognised by translocases in the mitochondrial membrane. In this review, we compare targeting sequences of different parasitic systems with highly reduced mitochondria and give an overview of how the import machinery has been modified -

Sulfur Cluster Synthesis in Toxoplasma
bioRxiv preprint doi: https://doi.org/10.1101/2021.01.28.428257; this version posted January 28, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Differential contribution of two organelles of endosymbiotic origin to iron- 2 sulfur cluster synthesis in Toxoplasma 3 Sarah Pamukcu1, Aude Cerutti1, Sonia Hem2, Valérie Rofidal2, Sébastien Besteiro3* 4 5 1LPHI, Univ Montpellier, CNRS, Montpellier, France 6 2BPMP, Univ Montpellier, CNRS, INRAE, Institut Agro, Montpellier, France 7 3LPHI, Univ Montpellier, CNRS, INSERM, Montpellier, France 8 * [email protected] 9 10 11 Abstract 12 Iron-sulfur (Fe-S) clusters are one of the most ancient and ubiquitous prosthetic groups, and they are 13 required by a variety of proteins involved in important metabolic processes. Apicomplexan parasites 14 have inherited different plastidic and mitochondrial Fe-S clusters biosynthesis pathways through 15 endosymbiosis. We have investigated the relative contributions of these pathways to the fitness of 16 Toxoplasma gondii, an apicomplexan parasite causing disease in humans, by generating specific 17 mutants. Phenotypic analysis and quantitative proteomics allowed us to highlight striking differences 18 in these mutants. Both Fe-S cluster synthesis pathways are necessary for optimal parasite growth in 19 vitro, but their disruption leads to markedly different fates: impairment of the plastidic pathway 20 leads to a loss of the organelle and to parasite death, while disruption of the mitochondrial pathway 21 trigger differentiation into a stress resistance stage. -

Genomes and Genome Projects of Protozoan Parasites
Curr. Issues Mol. Biol. (2003) 5: 61-74. Genomes of Protozoan Parasites 61 Genomes and Genome Projects of Protozoan Parasites Klaus Ersfeld other projects (Table 1) are at various stages of progress, but the reasoning and motivations for genomic research is School of Biological Sciences, 2.205 Stopford Building, virtually identical between all of them (Tarleton and University of Manchester, Oxford Road, Manchester M13 Kissinger, 2001). 9PT, UK. The Plasmodium falciparum genome Abstract More than 1 billion people are estimated to carry malaria- causing parasites at any one time. The annual mortality Protozoan parasites are causing some of the most rate is between 0.5-3 million people. A massive increase devastating diseases world-wide. It has now been in population, a deterioration in public health services and recognised that a major effort is needed to be able to infrastructures and the problems associated with now control or eliminate these diseases. Genome projects widespread drug resistance has led to a re-emergence of for the most important protozoan parasites have been malaria as one of the most serious diseases world-wide initiated in the hope that the read-out of these projects (http://www.who.int/tdr/diseases/malaria/default.htm). After will help to understand the biology of the parasites a period of optimism, mainly due to the relatively successful and identify new targets for urgently needed drugs. reduction of the malaria-transmitting mosquitoes with the Here, I will review the current status of protozoan pesticide DDT, more people die know of the disease than parasite genome projects, present findings obtained 40 years ago (Guerin et al., 2002a; Miller and Greenwood, as a result of the availability of genomic data and 2002). -

Why Chloroplasts and Mitochondria Retain Their Own Genomes and Genetic Systems
PAPER Why chloroplasts and mitochondria retain their own COLLOQUIUM genomes and genetic systems: Colocation for redox regulation of gene expression John F. Allen1 Research Department of Genetics, Evolution and Environment, University College London, London WC1E 6BT, United Kingdom Edited by Patrick J. Keeling, University of British Columbia, Vancouver, BC, Canada, and accepted by the Editorial Board April 26, 2015 (received for review January 1, 2015) Chloroplasts and mitochondria are subcellular bioenergetic organ- control. Fig. 2B illustrates the two possible pathways of synthesis elles with their own genomes and genetic systems. DNA replica- of each of the three token proteins, A, B, and C. Synthesis may tion and transmission to daughter organelles produces cytoplasmic begin with transcription of genes in the endosymbiont or of gene inheritance of characters associated with primary events in photo- copies acquired by the host. CoRR proposes that gene location synthesis and respiration. The prokaryotic ancestors of chloroplasts by itself has no structural or functional consequence for the and mitochondria were endosymbionts whose genes became mature form of any protein whereas natural selection never- copied to the genomes of their cellular hosts. These copies gave theless operates to determine which of the two copies is retained. Selection favors continuity of redox regulation of gene A, and rise to nuclear chromosomal genes that encode cytosolic proteins ’ and precursor proteins that are synthesized in the cytosol for import this regulation is sufficient to render the host s unregulated copy into the organelle into which the endosymbiont evolved. What redundant. In contrast, there is a selective advantage to location of genes B and C in the genome of the host (5), and thus it is the accounts for the retention of genes for the complete synthesis endosymbiont copies of B and C that become redundant and are within chloroplasts and mitochondria of a tiny minority of their lost. -

Apicoplast Fatty Acid Synthesis Is Essential for Pellicle Formation at the End of Cytokinesis in Toxoplasma Gondii Érica S
© 2016. Published by The Company of Biologists Ltd | Journal of Cell Science (2016) 129, 3320-3331 doi:10.1242/jcs.185223 RESEARCH ARTICLE Apicoplast fatty acid synthesis is essential for pellicle formation at the end of cytokinesis in Toxoplasma gondii Érica S. Martins-Duarte1,2,*, Maira Carias1,2, Rossiane Vommaro1,2, Namita Surolia3 and Wanderley de Souza1,2,* ABSTRACT an ‘apicoplast’, a non-photosynthetic secondary plastid originating The apicomplexan protozoan Toxoplasma gondii, the causative agent from a red algae plastid that was engulfed by an apicomplexan of toxoplasmosis, harbors an apicoplast, a plastid-like organelle with ancestor (Williamson et al., 1994; Köhler et al., 1997). Besides the essential metabolic functions. Although the FASII fatty acid evolutionary importance of the apicoplast, its discovery brought biosynthesis pathway located in the apicoplast is essential for new opportunities for the development of novel therapy targeting parasite survival, the cellular effects of FASII disruption in T. gondii diseases caused by apicomplexan parasites (Wiesner et al., 2008; had not been examined in detail. Here, we combined light and electron Goodman and McFadden, 2013). As in plant plastids, the apicoplast microscopy techniques – including focused ion beam scanning contains its own genome and pathways for the synthesis of electron microscopy (FIB-SEM) – to characterize the effect of FASII isoprenoids, fatty acids and iron-sulfur (Fe-S) clusters (Sheiner disruption in T. gondii, by treatment with the FASII inhibitor triclosan or et al., 2013; van Dooren and Striepen, 2013). More importantly, by inducible knockdown of the FASII component acyl carrier protein. owing to the prokaryotic ancestry of the apicoplast, its pathways are Morphological analyses showed that FASII disruption prevented divergent from the eukaryotic counterparts found in the mammalian cytokinesis completion in T.